bei meinen Beobachtungen von Bauchhärlingen in Mikroaquarien (https://www.mikroskopie-forum.de/index.php?topic=33985.0) habe ich immer wieder die Gelegenheit, die Entwicklung der Tiere über einige Wochen zu verfolgen. Da Gastrotrichen einen interessanten und im Tierreich seltenen - wenn nicht einzigartigen - Lebenszyklus besitzen, möchte ich Euch einladen, mit mir den Lebensweg des Bauchhärlings Chaetonotus brevisetosus zu begleiten.
Ch. brevisetosus ROSZCZAK 1935 ist ein mit ca. 210µm Länge ein recht stattlicher Gastrotrich, der sich im Schuppenkleid von Ch. polyspinosus praktisch nicht unterscheidet. Da die deutlich kleinere Länge von Ch. brevisetosus kein gutes Kriterium zur Artabgrenzung ist und die Artfestlegung bei Bauchhärlingen traditionell durch die Schuppen erfolgt, sehen einige Bearbeiter Ch. brevisetosus als ein Synonym zu Ch. polyspinosus an. Eine längere Beobachtung in einem Mikroaquarium enthüllt aber einige Unterschiede zu Ch. polyspinosus (Magen, Form des X-Organs, Eiform, Verhalten).
Die Festlegung der Artgrenzen ist bei (zumindest vorwiegend) parthenogenetischen Tieren sehr problematisch. Die klassische biologische Artdefinition als Fortpflanzungsgemeinschaft versagt in diesem Fall und es verbleibt nur eine morphologische Beschreibung der Arten. Die Unterschiede zwischen zwei Individuen ist dann aber von der Genauigkeit der Beobachtung abhängig und erscheint daher oft willkürlich und nur der für die Klassifizierung in der praktische Arbeit notwendig.
Da sowohl Ch. polyspinosus als auch Ch. brevisetosus bei mir im selben Habitat vorkommen und deutlich morphologisch und durch ihr Verhalten zu unterscheiden sind, verwende ich im folgenden weiterhin die Bezeichnung Ch. brevisetosus. Ob diese Tiere eine eigene Art oder eine Variante von Ch. polyspinosus darstellen, bleibt der Weisheit der Taxonomen vorbehalten. Da Ch. brevisetosus seit seiner Erstbeschreibung im polnischen Tiefland wohl nicht mehr gesichtet wurde, werde ich im folgendem auf die Unterschiede zu Ch. polyspinosus hinweisen.
Wie bei allen Gastrotrichen beginnt das Leben von Ch. brevisetosus als Ei. Die Eier werden einzeln und offen abgelegt und haben eine doppelte, durch kleine - bei Wasserkontakt aufquellenden - Säulen zwischen den Eischalen versteifte Schale. Typisch für Bauchhärlinge sind die kristallinen Körper in den Darmzellen der Embryos, deren Funktion unbekannt ist und die sich in den ersten Lebensstunden auflösen. Zum Vergleich ist ein Bild eines Eies von Ch. polyspinosus aus dem selben Habitat eingefügt.
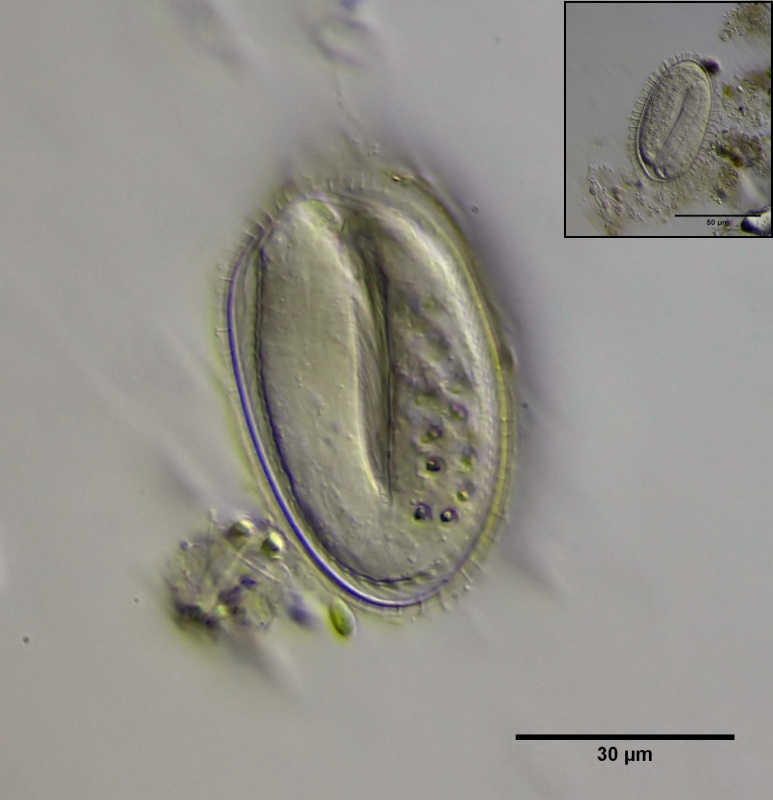
Bild 1: Ch. brevisetosus, Ei kurz vor dem Schlupf; Inset: Ei von Ch. polyspinosus zum Vergleich
Kurz nach dem Schlupf zeigen sich die für Jungtiere typischen Proportionen. Kopf und Saugmagen (Pharynx) sind bereits voll ausgewachsen, während der Hinterleib mit dem Darm noch deutlich verkürzt ist. Das anschließende Wachstum des Tieres erfolgt daher nur am Hinterleib, bis die Erwachsenengröße nach einigen Tagen erreicht wird.
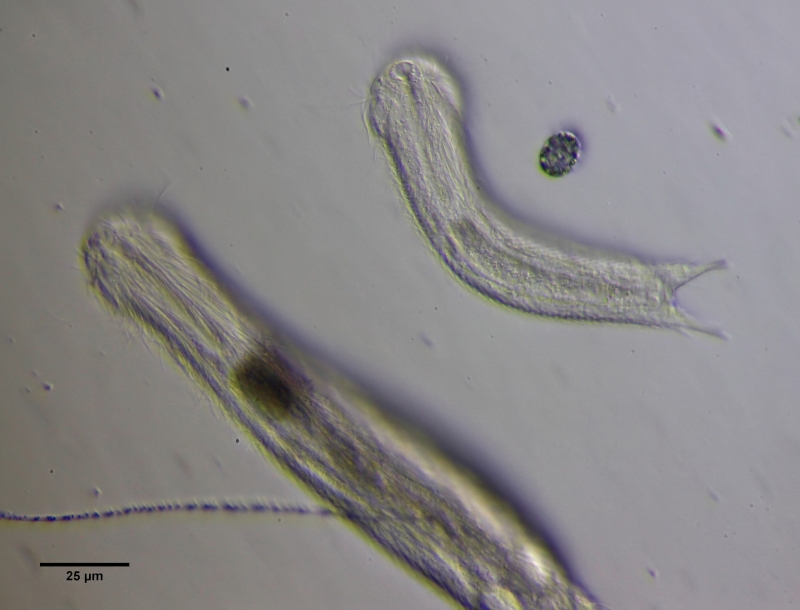
Bild 2: Ch. brevisetosus, Größenvergleich Jungtier und ausgewachsenes Tier
Bereits kurz nach dem Schlupf ist am Darmeingang ein nahezu ungefärbter Bereich sichtbar. Dieser - später goldbraun gefärbte - Bereich wird als "Magen" bezeichnet und tritt nur bei sehr wenigen Gastrotrichen-Arten auf und bildet ein weiteres Unterscheidungskriterium zu Ch. polyspinosus der einen solchen Magen nicht besitzt.

Bild 3: Ch. brevisetosus, kurz nach dem Schlupf; am Darmeingang ist die noch ungefärbte Anlage des "Magens" zu erkennen
Im Laufe der folgenden Entwicklung färbt sich der Magen des Tieres zunehmend.

Bild 4: Ch. brevisetosus, älteres Jungtier mit leicht gefärbten Magen
Da der Magen anfänglich nur schwach gefärbt ist, kann seine Struktur in unterschiedlichen Schärfeebenen gut als Ring um den Darmeingang erfasst werden.
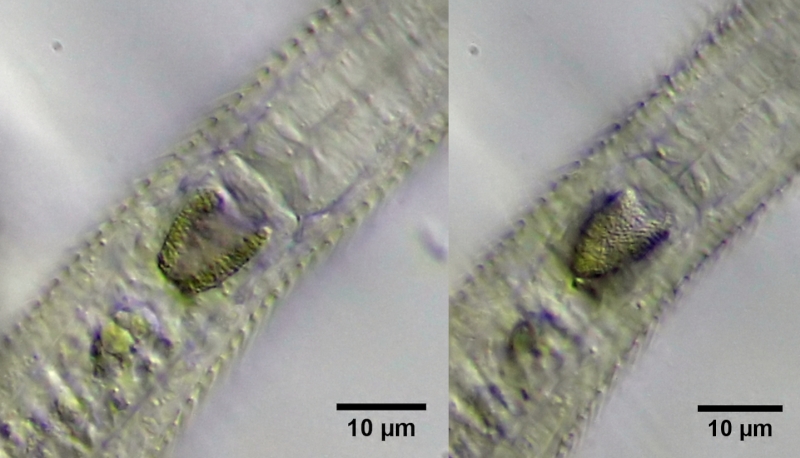
Bild 5: Ch. brevisetosus, unterschiedliche Fokuslagen durch den noch leicht gefärbten Magenring
Bei älteren Tieren ist der Magen praktisch undurchsichtig und erscheint als eine dunkle, granuläre Masse. Die Funktion des Magens ist unbekannt. Da der Magen anfänglich ungefärbt ist, gehe ich davon aus, dass es sich hierbei um ein Speicherorgan handelt, in dem Nahrungsreserven eingelagert werden. Ich habe bisher nur zwei Gastrotrichenarten mit Magen gefunden (Heterolepiderma majus und eben Ch. brevisetosus). Beide Arten zeigen ein für Bauchhärlinge sehr auffälliges Verhalten: Gastrotrichen sind normalerweise immer auf Nahrungssuche und nie in Ruhe anzutreffen. Lediglich bei den beiden "Magen-Arten" fällt auf, dass die Tiere auch mal minutenlang an einer Stelle verharren und die Nahrungssuche unterbrechen. Daher gehe ich davon aus, dass die Existenz eines Magens die Effektivität des Verdauungssystems erhöht und eine ständige Nahrungsaufnahme nicht mehr nötig ist.
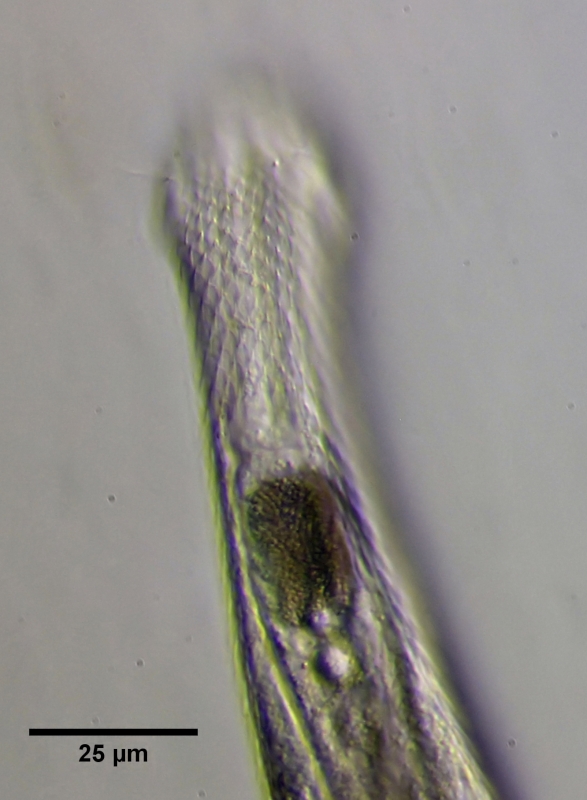
Bild 6: Ch. brevisetosus, erwachsenes Tier, Magen
Nach einigen Tagen sind die Tiere ausgewachsen und treten in die stressige parthenogenetische Reproduktionsphase ein.
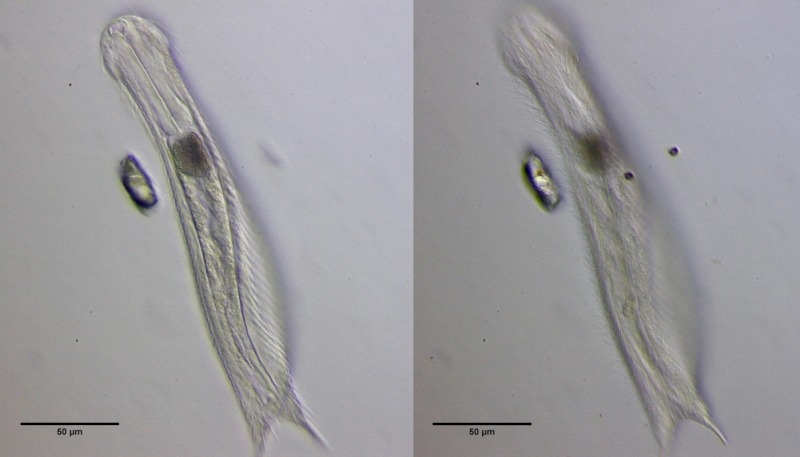
Bild 7: Ch. brevisetosus, erwachsenes Tier, links mittiger Schnitt, rechts Fokus auf die Beschuppung der Bauchseite
Die dorsalen Schuppen des Tieres sind sehr kleinteilig und erst nach einer Schuppenanalyse ist die pfeilfömige Gestalt zu erkennen. Schuppenform und (dorsale) Schuppenveilung sind nicht von Ch. polyspinosus zu unterscheiden.

Bild 8: Ch. brevisetosus, erwachsenes Tier, dorsales Schuppenkleid; Inset: abgelöste Einzelschuppe
Ein Fokus auf die Kopfunterseite macht deutlich, woher der Name "Bauchhärlinge" abgeleitet ist: Beidseitig zieht sich je ein Band von Zilien entlang, die der Fortbewegung des Tieres dienen. Gut zu sehen sind die Basalkörper der Einzelzilien. Bei Ch. brevisetosus sind - anders als bei Ch. polyspinosus - die beiden Zilienbänder hinter der Mundöffnung verbunden. Außerdem fehlt eine ausgeprägte kutikulare Platte direkt hinter dem Mund (das "Hypostomium") das bei Ch. polyspinosus prominent ausgebildet ist und zwei Höcker trägt.
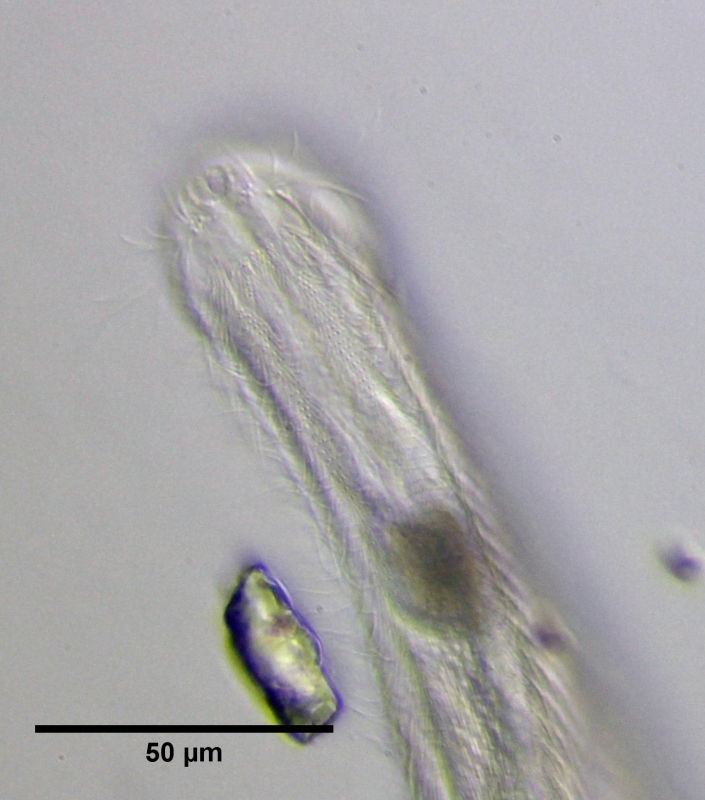
Bild 9: Ch. brevisetosus, erwachsenes Tier, ventrale Zilienbänder
In dieser parthenogenetischen Phase legen die Tiere (meist) vier unbefruchtete, im Vergleich zur Größe der Tier, riesige Eier im Abstand von ca. einem Tag. Anders als bei der "Laborratte" der Gastrotrichen-Forscher Lepidodermella squammata werden keine Dauereier abgelegt. Alle Eier entwickeln sich sofort zu Embryos weiter. Eine der häufigsten Todesursachen bei Gastrotrichen ist, dass die riesigen Eier oft nicht abgelegt werden können und die Mutter nach einem stundenlangen Kampf verstirbt. Diese enorm anstrengende parthenogenetische Reproduktionsphase schwächt die Tiere so, dass auch Krankheiten gehäuft auftreten wie zum Beispiel der in Bild 10 gezeigte bakterielle Befall des Darmes durch Bakterien. Das Tier konnte keine Nahrung mehr aufnehmen und verstarb bald nach der Aufnahme. Beachtenswert finde ich, das der Magen bei dem befallenen Tier wieder ungefärbt ist. Möglicherweise wurden die Vorratsstoffe, die den Magen färben, aufgebraucht.
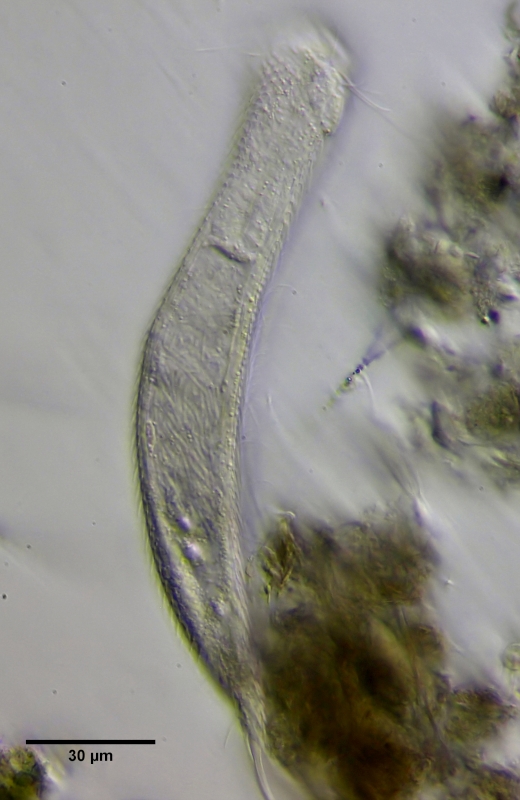
Bild 10: Ch. brevisetosus, krankes Tier mit "Daarmentzündung"
Die Sterblichkeit in dieser Entwicklungsphase ist deshalb sehr hoch und die durchschnittliche Lebenserwartung der Tiere beträgt nur ein bis zwei Wochen. Oft findet man verstorbene Tiere im Präparat, von denen meist nur noch die kutikularen Teile zu erkennen sind. Dies erlaubt es, das Schuppenkleid der Tiere genauer zu studieren.

Bild 11: Ch. brevisetosus, verstorbene Tiere
So ist es auch in einem Mikroaquarium möglich, genau Bilder von den ventralen Terminalplatten zu gewinnen, die oft für die Artbestimmung ausschlaggebend sind. Bei Ch. brevisetosus sind - anders als bei Ch polyspinosus - zwei große Terminalplatten vorhanden.

Bild 12: Ch. brevisetosus, Terminalplatten
Bei den wenigen Überlebenden dieser parthenogenetischen Phase beginnt ein Umbau des Tieres zu einem Zwitter. In dieser "post-parthenogenetischen" Phase werden keine Eier mehr abgelegt und die Lebenszeit des Tieres verlängert sich um bis zu drei weiteren Wochen.
Der Umbau zum Zwitter beginnt mit der Entwicklung des X-Organs am Hinterende, beidseitig unterhalb des Darms des Tieres.
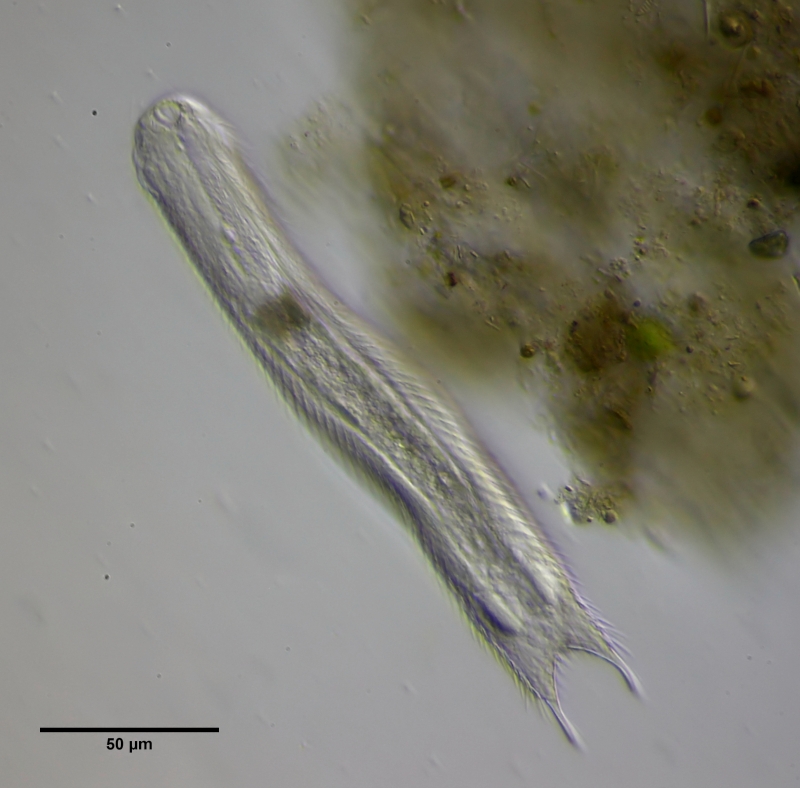
Bild 13: Ch. brevisetosus, Beginn der Entwicklung des X-Organs
Einige Zellen der Ovarien beginnen sich zu teilen und schließen sich zu beidseitig zu je einer Drüse zusammen, die in einen inneren Hohlraum ein Sekret mit unbekannter Funktion abgeben. Am Ende dieser Entwicklung ist deutlich die Hülle des X-Organs und das Sekret im Inneres des Organs zu unterscheiden. Die beiden Sphären des X-Organs sind miteinander verbunden.
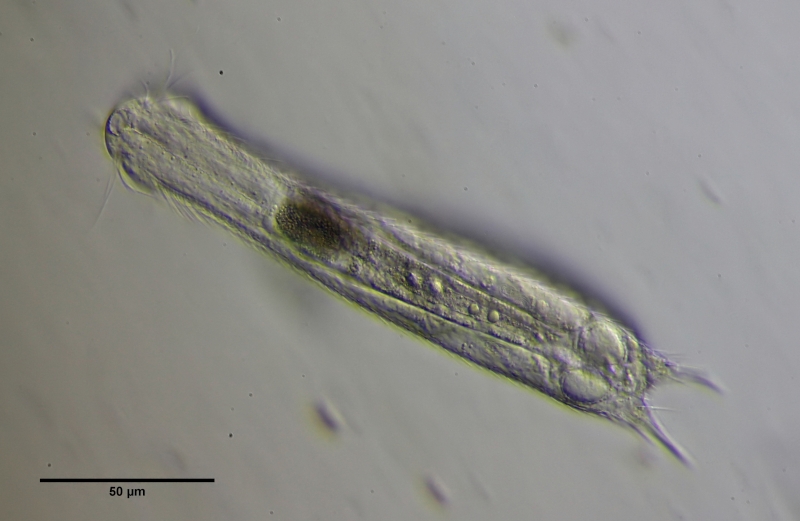
Bild 14: Ch. brevisetosus, voll entwickeltes X-Organ
In der Seitenansicht erkennt man die Lage des X-Organs und die Trennung zwischen granuliertem Cortex und dem homogenen Inhalt.
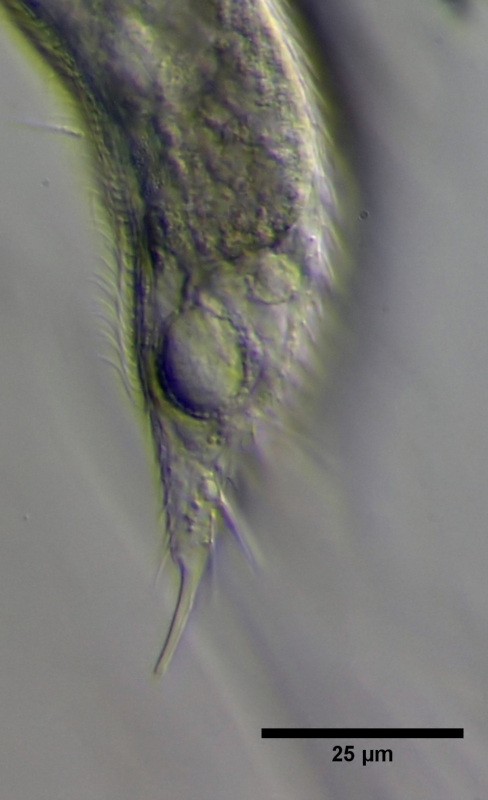
Bild 15: Ch. brevisetosus, Seitenansicht des X-Organs
Bei Ch. brevisetosus ist gut zu erkennen, dass das X-Organ einen mittigen Ausführungsgang zur Bauchseite des Tieres besitzt, der unter den Schuppen endetet.

Bild 16: Ch. brevisetuosus, Ausführungsgang des X-Organs
Parallel zu dem X-Organ entwickeln sich nahe der Bauchseite einige unbewegliche Spermapakete, die im Laufe der Entwicklung weiter zum Hinterende des Tieres an das Ende des sich parallel entwickelnden Eies wandern. In der Endlage liegen die Spermapakete in der Nähe Stelle, an der auch die Eier ausgeschieden werden (die Gonopore ist mikroskopisch wohl nicht zu erkennen).
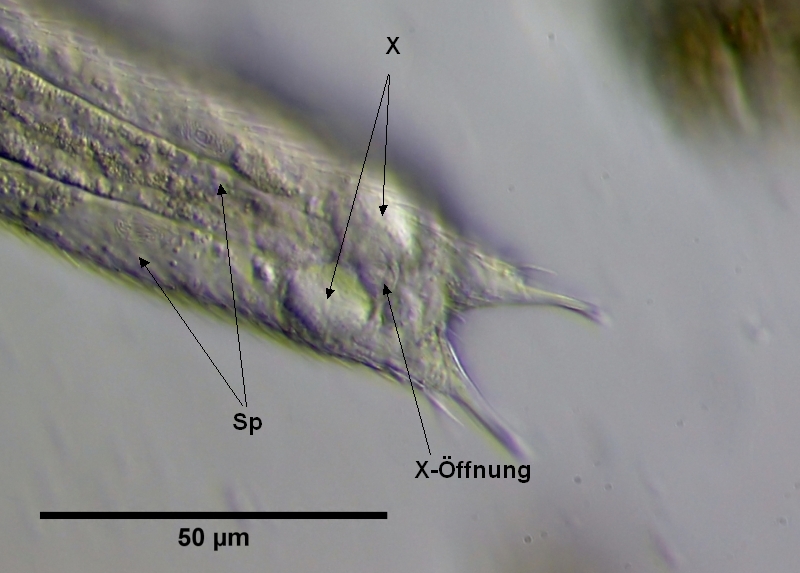
Bild 17: Ch. brevisetosus, Spermapakete; Sp: Spermapakete, X: X-Organ; X-Öffnung: Ventrale Öffnung des X-Organs
Fokussiert man in der Seitenansicht auf die Mitte des Tieres, ist die Lage des Ausführungsgangs des X-Organ gut zu erkennen. Zusätzlich zu den Spermapaketen ist ein reifes Ei voll entwickelt. Das Tier ist zu einem echten Zwitter konvertiert. Zu diesem Zeitpunkt ändert sich das Verhalten des Tieres. Normalerweise bilden die meisten Gastrotrichenarten Kleingruppen. Findet man in einem Mikroaquarium ein Tier, halten sich auch die anderen Tiere in der Nähe auf. Die post-parthenogenetischen Zwittertiere sondern sich von dieser Gruppe ab. Möglicherweise versucht das Tier nun, sich anderen Kleingruppen anzuschließen um zur Paarung die genetische Vielfalt zu erhöhen.
Innerhalb eines Mikroaquariums konnte keine Paarung oder Eiablage eines Zwittertiers beobachtet werden. Das Tier überlebte ca. drei Wochen nach der Umwandlung zu einem Zwitter bevor es verstarb.
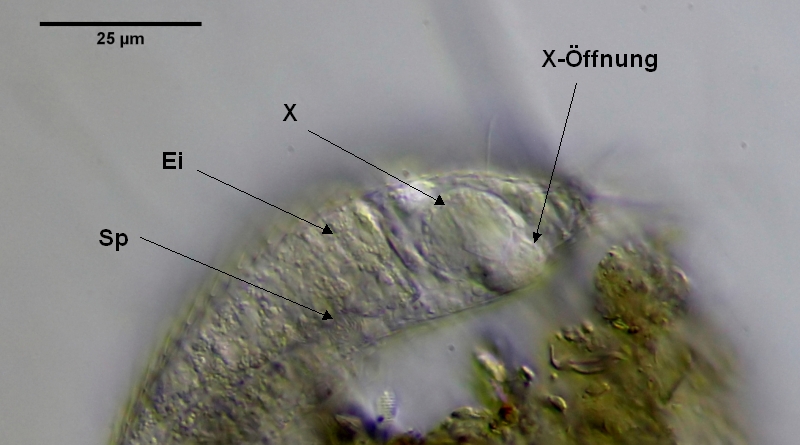
Bild 18: Ch. brevisetosus, Seitenansicht; Ei: reife Eizelle; Sp: Spermapakete, X: X-Organ; X-Öffnung: Ventrale Öffnung des X-Organs
Am Ende der Lebensdauer des Mikroaquariums verschlechtern sich die Lebensbedingungen für die Tiere mehr und mehr und die aktiven Tiere sterben. Die zu diesem Zeitpunkt abgelegten Eier verlangsamen ihre Entwicklung immer mehr, bis die voll entwickelten Embryos scheinbar leblos in den Eiern liegen. Erst eine Zeitrafferaufnahme der Eier enthüllt, dass die Tiere (auch nach einigen Monaten) immer noch leben und sich ganz langsam bewegen. Ähnliches hatte ich schon mal für Lepidochaeta zelinkai dargestellt (https://www.mikroskopie-forum.de/index.php?topic=31200.0). Ich gehe davon aus, dass Ch. brevisetosus keine Dauereier bildet, sondern bei "schlechte Zeiten" den Schlupf zurückstellt und die Embryonalentwicklung unterbricht. Verbessern sich die Verhältnisse wieder, schlüpft die neue Generation aus den überdauerten Eiern.
Ich hoffe, Euch auf eine interessante Reise ins Reich der Gastrotrichen mitgenommen zu haben.
Viele Grüße
Michael