Eine der erstaunlichsten Aspekte bei der Entwicklung von Organismen, ist in meinen Augen die Bildung von unbelebten Strukturen. Es gibt fast kein Lebewesen, das ohne artspezifische unbelebte Strukturen auskommt - seien es Knochen, Krallen und Haare bei Wirbeltieren, das Außenskelett der Insekten oder die Zellwände von Pflanzen. Wie können diese hochspezialisierten und artspezifischen Strukturen innerhalb und außerhalb von Zellen entstehen, wo doch nur die Zellen der Träger der genetischen Informationen sind? Ich möchte in diesem Beitrag die Bildung von unbelebten Strukturen anhand von Zeitraffer-Aufnahmen der Embryonalentwicklung einiger typischer Wasserbewohnern genauer vorstellen. Alle Aufnahmen entstanden in Mikroaquarien. Für den Celops- und den Gastrotrichenfilm wurde das 100er Objektiv verwendet, für den Rädertierfilm ein 50er Wasserimmersionsobjektiv. Um die Entwicklungen deutlich zu machen, mussten die Filme zum Teil stark beschleunigt werden (Coleps 10x, Lecane 800x, Ch. cordiformis 400x).
1. Bildung der Panzers bei Coleps hirtus nach einer Teilung
Das häufige Wimperntier Coleps hirtus mit seinem auffälligem Panzer ist wohl den meisten von Euch bekannt.

Bild 1: Coleps hirtus; Panzer
Der Panzer besteht aus einer Reihe von Einzelplatten, die innerhalb der Zelle, in Vakuolen des Zellcortex liegen.
Wenn sich der Ciliat teilt, wird der Panzer des Muttertieres gerecht zwischen den beiden Tochtertieren aufgeteilt. Die jeweils fehlende Hälfte des Panzers muss dann von den beiden Tochterzellen nachgebildet werden.

Bild 2: Coleps hirtus; Tochtertiere gegen Ende der Teilung; die Pfeile zeigen auf Vesikel, die Material zur Panzerbildung transportieren
Bereits während der Teilung - vor Trennung der Tochterzellen - beginnt die Bildung der neuen Panzerplatten. An den Grenzen der neuen Platten wird von Vesikeln Material abgelagert und die Konturen der neuen Panzerplatten sind bereits zu erahnen. Elektronenmikroskopische Untersuchung 1 zeigen, dass diese Vesikel vor allem Calcium beinhalten, das als amorphes Calciumcarbonat in die Platten eingebaut wird.

Bild 3: Coleps hirtus; fortschreitende Panzerbildung
In den folgenden Stunden wird immer mehr Material abgelagert und die Panzerstruktur immer deutlicher sichtbar.
Das Material für die Panzerplatten wird an einer genetisch vorgebildeten Matrix innerhalb der Zellen abgeschieden. Diese Matrix (deren Bildung im Detail noch nicht verstanden ist) markiert also die Orte, an denen die Vesikel ihr Material deponieren und so die fein strukturieren, komplex geformten Panzerplatten langsam aufbauen.
Den gesamten Prozess zeigt folgender Zeitrafferfilm zusammengefasst innerhalb von 2:30 min, der mit einem "Click" auf das folgende Bild gestartet werden kann

Bild 4: Coleps hirtus Zeirafferfilm zur Panzerbildung
(Zum Starten bitte anklicken!)
Innerhalb von Zellen funktioniert also die Bildung unbelebter Strukturen meiner Meinung nach folgendermaßen:
- Die Zellen bilden genetisch gesteuert eine Art Matrix, die die Position und Form der unbelebten Strukturen festlegt
- Die Zellen produzieren das Material der Strukturen, das in Vesikel verpackt wird.
- Die Vesikel werden zur Matrix transportiert und lagern dort das Baumaterial der Strukturen ab.
Viele unbelebte Strukturen befinden sich aber nicht innerhalb von Zellen. Ein Beispiel dafür sind die Trophi (die Hartteile des Kauers) von Rädertieren, die außerhalb der Mastaxzellen in einem Hohlraum gebildet werden.
2. Bildung der Hartteile des Kauers (Trophi) bei Lecane sp.
Kauer von Rädertieren setzt sich aus einer Reihe von artspezifischen Hartteilen (Trophi) zusammen, die während der Embryonalentwicklung innerhalb des Eis aus Chitin gebildet werden. Diese Strukturen befinden sich im Lumen des Mastax der Tiere, also außerhalb der Zellen. Wie steuern also die Zellen - als Träger der genetischen Informationen - die artspezifische Form der Trophi?
Beobachtet man den Verlauf der Embryonalentwicklung (hier am Beispiel einer Lecane-Art), kann man den Ablauf der Strukturbildung beobachten.
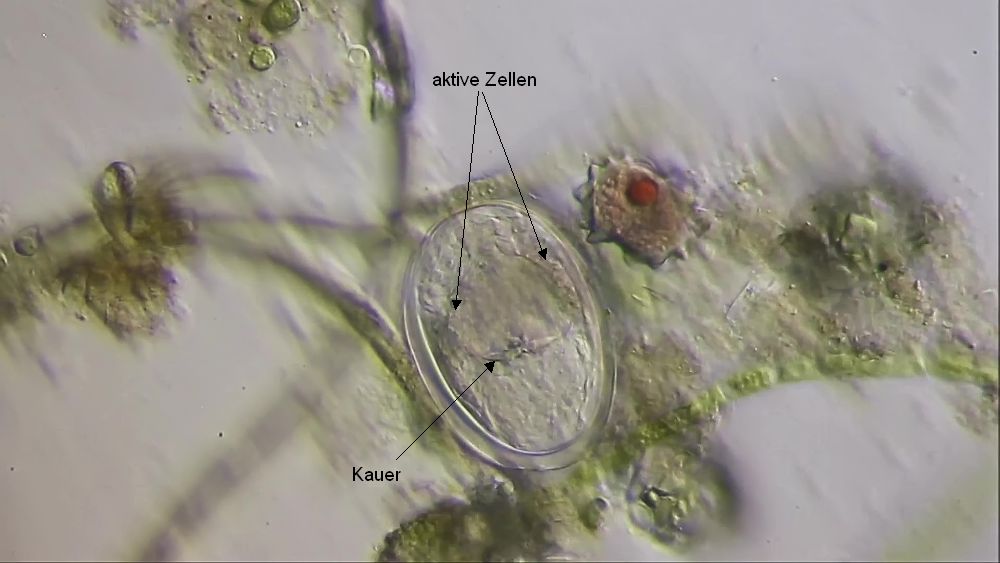
Bild 5: Lecana sp.; Bildung der Trophi
Die Bildung der Trophi erfolgt an der Wand des Mastax. Sie werden - wie die Panzerplatten bei Coleps - über Ihre gesamte Ausdehnung gleichmäßig abgeschieden. Das Material für die Bildung wird in einem Ring aktiver Zellen in der Mastax-Wand produziert und an die Zellwand transportiert. Hier wird das Material an der Zellwand entlang einer zellübergreifenden, genetisch bestimmten Matrix auf der Zellwand außerhalb der Zellen abgeschieden.

Bild 6: Lecane sp. Zeitliche Abfolge der Trophibildung
Die einzelnen Trophipaare werden nacheinander gebildet (Reihenfolge: Rami, Unci, Manubria - Fulcrum lag leider außerhalb der Fokusebene). Die von den Zellen vorgegebene Strukturinformation wird also während der Embryonalentwicklung angepasst, so dass eine zeitlich abgestimmte Bildung der Trophi erfolgt.
Der Gesamtprozess ist in folgendem Video (1:10 min) in 800-facher Geschwindigkeit zu sehen:
Bild 7: Lecane sp.; Zeitrafferfilm zur Trophi-Bildung
(Zum Starten bitte anklicken!)
Außerhalb, aber in der Nähe von Zellen innerhalb des Körpers funktioniert also die Bildung unbelebter Strukturen meiner Meinung nach folgendermaßen:
- Die Zellen bilden nahe ihrer Oberfläche genetisch gesteuert eine Art Matrix, die die Position und Form der unbelebten Strukturen festlegt
- Die Zellen Produzieren das Material der Strukturen, das in Vesikel verpackt wird.
- Die Vesikel werden zur Matrix transportiert und lagern dort das Baumaterial der Strukturen ab.
Wie klappt aber die Strukturbildung weit entfernt von den Zellen außerhalb des Körpers von Tieren? Ein Beispiel für diese Verhältnisse sind die Schuppen und Stacheln von Gastrotrichen.
3. Bildung der Haftröhrchen, Cutikularplatten und Schuppen bei Chaetonotus cordiformis
Süsswasser-Gastrotrichen (genauer Chaetonotoidea) besitzen eine sehr aufwändig skulpturierte Cutikula. Die äußere Hülle wird von einer sehr dünnen, etwa 100nm dicken Epicutikula gebildet. Dieses dünne Häutchen bedeckt das gesamte Tier, einschließlich aller Cilien, Sinneshaare und Stachelschuppen und ist wohl ein einzigartiges Merkmal innerhalb des Tierreichs. Unter der Epicutikula liegen die sehr artspezifischen und meist aufwändig gestaltete Schuppen der Tiere. Diese Schuppen sind oft nur lose mit der nächsten, ca. 1µm dicken Cutikula-Schicht, der Endocuticula verbunden. Diese Endocuticula liegt direkt auf den Hautzellen der Tiere auf.
Zusätzlich besitzen diese Bauchhärlinge einige dickere Cutikula-Platten, die wohl Verdickungen der Endocutikula sind und von denen das Wachstum des Kopfschildes (Kephalion) im Film verfolgt werden kann. An den Zehen sitzen hohle Haftröhrchen, in die die Klebedrüsen ihre Sekrete abgeben und die zum blitzschnellen Festheften der Tiere an das Substrat dienen.
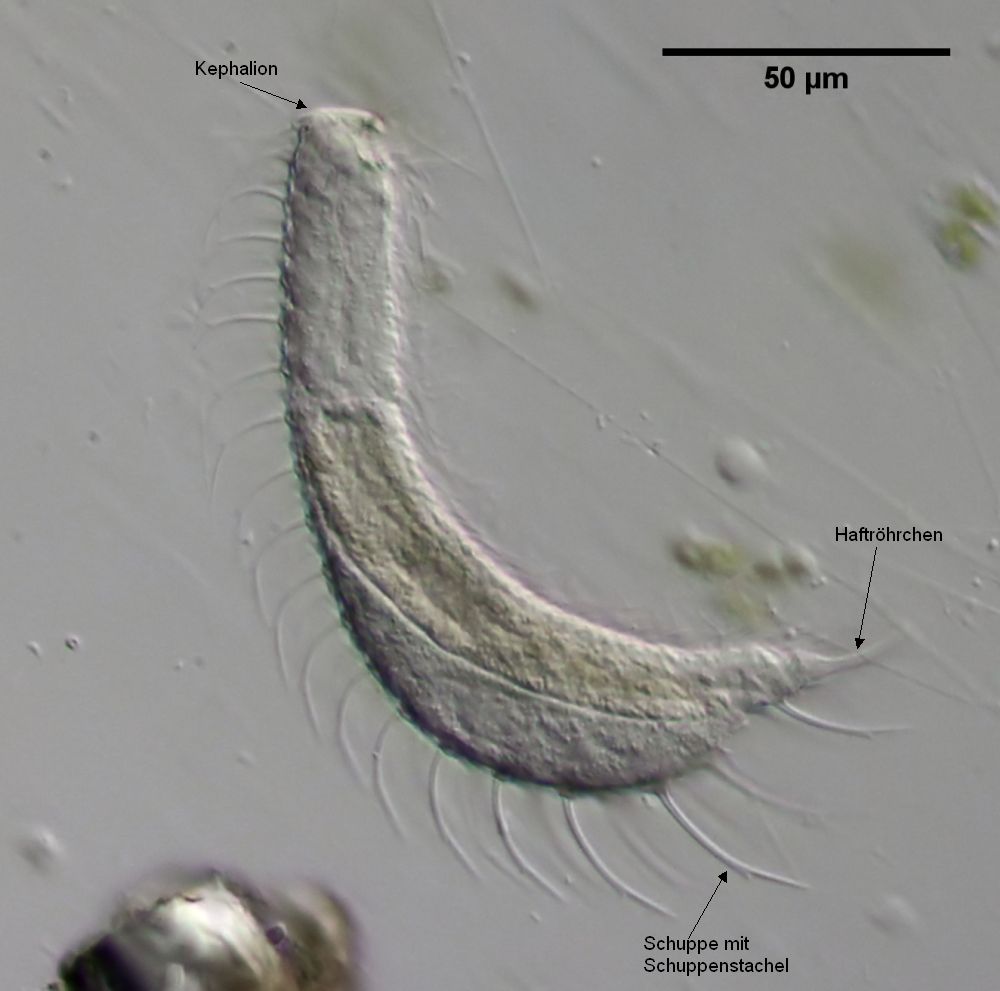
Bild 8: Ch. cordiformis; Cutikulare Strukturen
Bei Ch. cordiformis sind die Haftröhrchen relativ lang und laufen spitz zu. Auffällig ist das Kephalion, das bei dieser Art über den Rücken der Tiere hinausragt und dessen Ende charakteristisch gebogen ist.
Am aufwändigsten sind die herzförmigen - bis längsovalen - Schuppen der Tiere gestaltet, die einen sehr langen Stachel tragen, der im hinteren Drittel der Basisplatten entspringt und recht weit von seinem Ende entfernt eine auffällige Nebenspitze trägt.
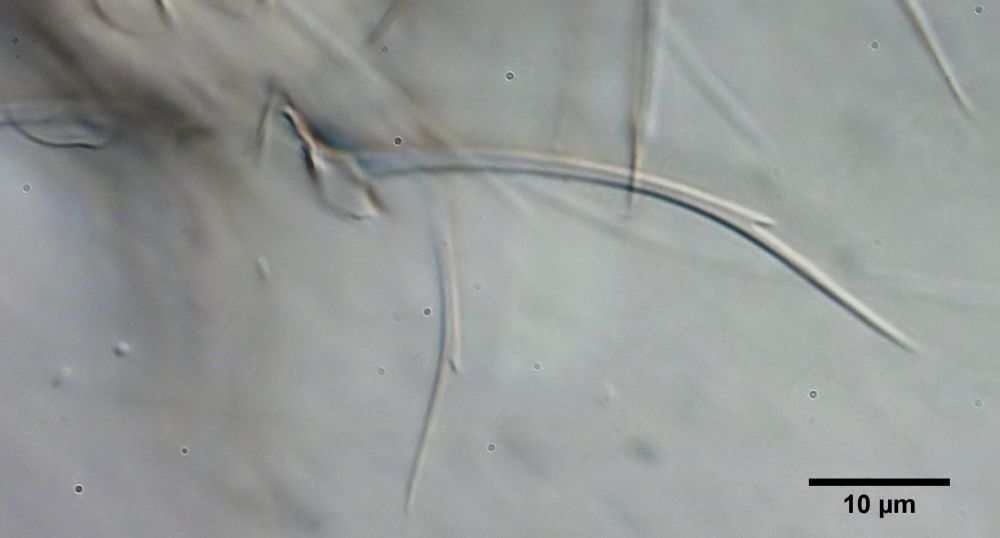
Bild 9: Ch. cordiformis; Stachelschuppe mit Nebenspitze
Wie werden diese unbelebten Strukturen so artspezifisch gebildet, obwohl sie sehr weit von den Hautzellen, die ja die genetische Information tragen, entfernt sind? Dazu muss man die Embryonalentwicklung der Tiere verfolgen.
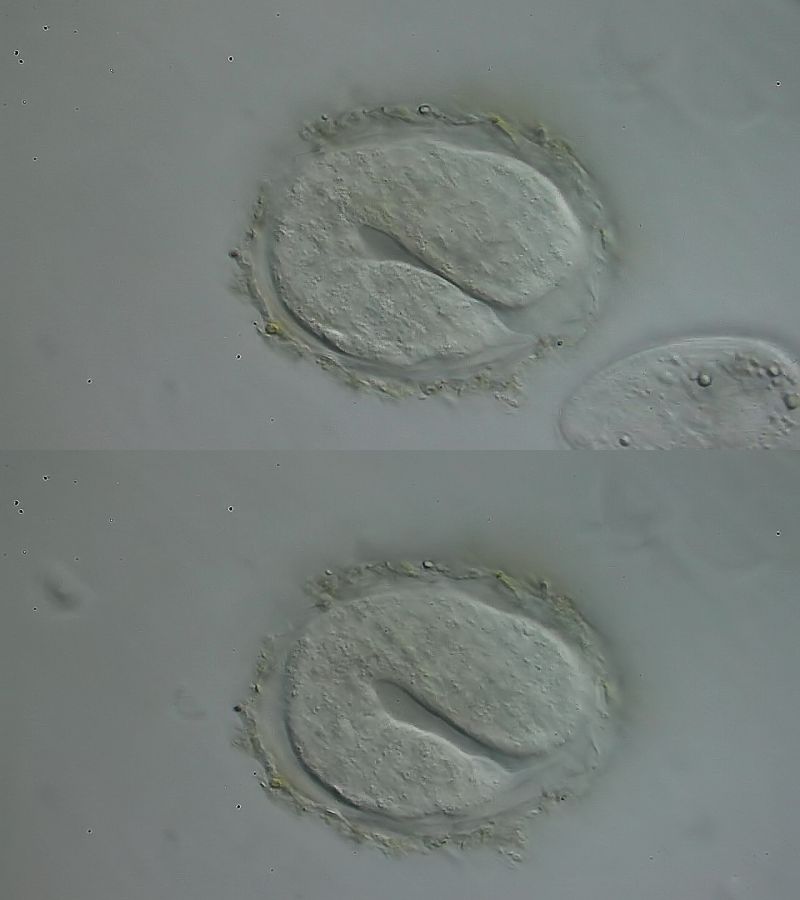
Bild 10: Ch. cordiformis; Bildung der Haftröhrchen
Fokussiert man nach dem Abschuss der Teilungsphase der Zellen auf die Hautoberfläche des Embryos innerhalb des Eis, erkennt man, dass auf den Hautzellen sehr viele Vesikel unterwegs sind, die wohl von den Hautzellen ausgeschieden werden. Sie transportieren das Material, das zur Bildung der Cutikula notwendig ist. Anders als bei den beiden obigen Beispielen werden die cutikularen Strukturen nicht über das gesamte Volumen gleichmäßig gebildet, sondern wachsen nur an der Kontaktfläche zu den Zellen. Hier wird das Material entlang einer genetisch vorgegeben, zeitlich variablen Matrix abgelagert. Deutlich erkennt man, dass z. B. die Haftröhrchen langsam in der Länge wachsen, bis die endgültige Gestalt erreicht ist.
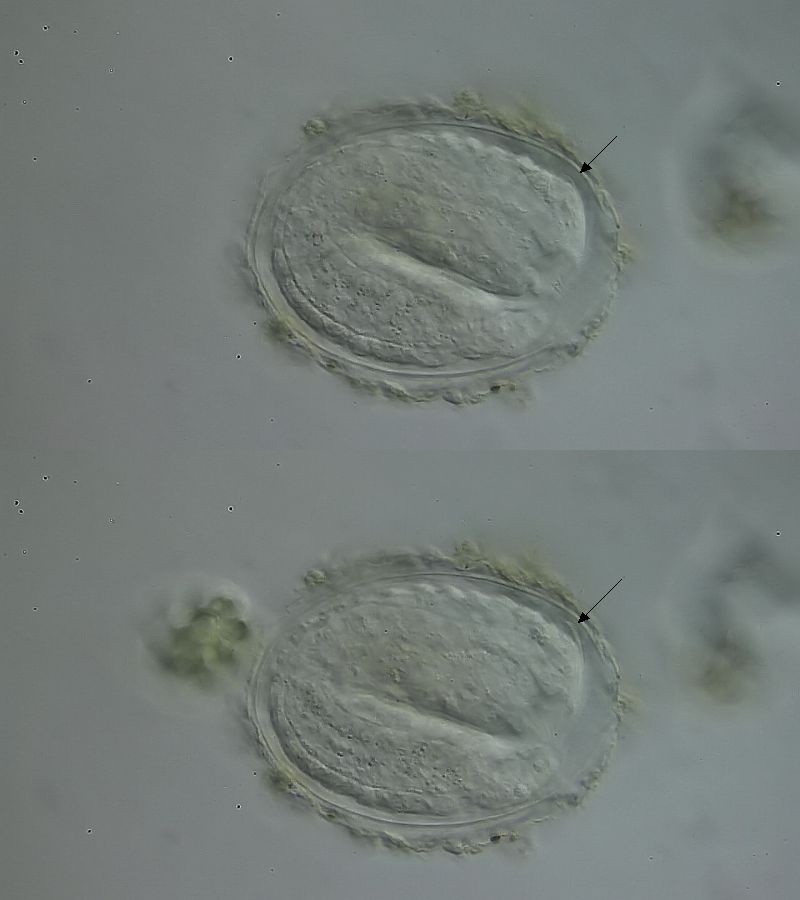
Bild 11: Ch. cordiformis; Wachstum des Kephalions
Besonders deutlich ist dieser Prozess bei dem Wachstum des Kephalions zu sehen. Das charakteristisch gebogene Ende des Kopfschildes wurde bereits zu Anfang des Wachstums in seiner endgültigen Form gebildet und wird im Laufe der Zeit immer weiter über den Rücken des Tieres hinausgeschoben.
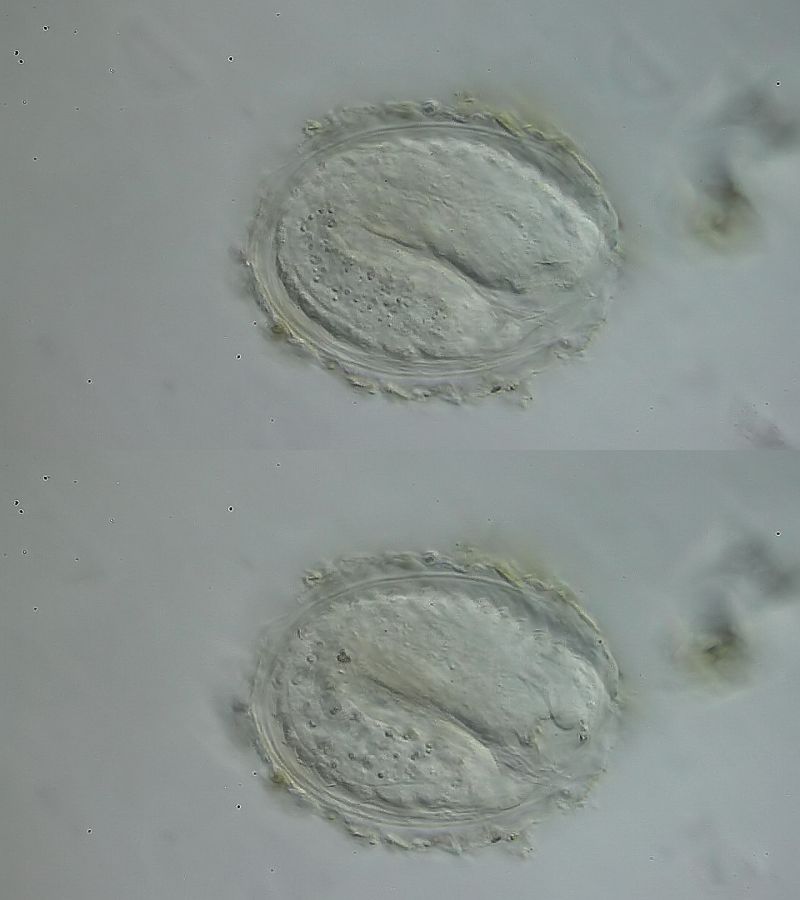
Bild 12: Ch. cordiformis; Bildung der Schuppen
Auch die Schuppenbildung verläuft analog. Zuerst werden die langen Stacheln gebildet, deren Struktur durch die zeitlich Modulation der formgebenden Matrix bestimmt wird. Erst am gegen Ende des Prozesses - nach der Bildung der langen Stacheln - werden die Basisplatten der Schuppen sichtbar. Dieser Bildungsprozess erinnert an einen biologischen 3D-Drucker: Schicht für Schicht wird das Material programmgesteuert abgelagert bis das endgültige Werkstück - z. B. die Stachelschuppen - vollständig gebildet sind.
Die Bildung der cutikularen Strukturen bei Chaetonotus cordiformis kann direkt in einem ca. 4 minütigem Zeitrafferfilm in 400 facher Geschwindigkeit verfolgt werden:
Bild 13: Ch. cordiformis; Zeitrafferfilm zur Cuticula-Bildung
(Zum Starten bitte anklicken!)
Die biologisch Musterbildung und speziell die Bildung der unbelebten Strukturen ist - soviel ich weiß - noch nicht endgültig verstanden und ein aktuellen Forschungsthema. Auch wenn wir Amateure hier wenig betragen können, lässt mich - und vielleicht auch Euch - die Beobachtung dieser Vorgängen staunend und fasziniert zurück.
Viele Grüße
Michael
--------------------------------
Literatur:
1 M. L. Lemloh et al., Genesis of amorphous calcium carbonate containing alveolar plates in the ciliate Coleps hirtus (Ciliophora, Prostomatea), J. Struct. Biol., vol. 181, no. 2, pp. 155–161, 2013, doi: 10.1016/j.jsb.2012.12.001.