als Max Voigt 1903 den Schlamm einiger Teiche im Schlosspark von Plön untersuchte, fand er eine sehr vielfältige Gastrotrichenfauna, die man damals im "Faulschlamm" (Sapropel) von Teichen nicht vermutet hätte. Einer dieser seltsamen, Schwefelwasserstoff-toleranten Bauchhärlinge aus diesem sauerstoffarmen Milieu war der neu entdeckte
Chaetonotus arquatus (VOIGT, 1903)
Da dieser selten gefundene und nicht sehr gut untersuchte Gastrotrich noch niemals fotografisch dokumentiert wurde, beschloss ich, diese Tiere hier vorzustellen, als ich das Glück hatte, eine stabile Population in einem nahe gelegenen Waldtümpel anzutreffen. Als ich begann, mich näher mit diesem seltsamen Tier zu beschäftigen, war mir noch nicht klar, wie einzigartig diese Art unter den Gastrotrichen ist...
Chaetonotus arquatus ist ein mittelgroßer (ca. 220µm) Bauchhärling, der sofort durch seine langen, gebogenen Stacheln auffällt.

Bild 1: Ch. arquatus; Querschnitt
Die markante, leicht S-förmig geschwungene Schwanzgabel macht das Tier unverwechselbar. Jeder Stachel entspringt einen napfförmigen Basischuppe, die einer Ausbuchtung des Körpers aufsitzt. Dadurch erinnert das gesamte Tier an ein "Michelin-Männchen".

Bild 2: Ch. arquatus; lateraler Schnitt
In der Seitenansicht offenbart sich diese eigenartige Körperform und die dazugehörigen Schuppen deutlicher. Die Schuppen selbst sind nicht durch die üblichen Methoden separierbar und stellen wohl eine Verdickung der Kutikula dar, aus der sich die hohlen (mit einem Septum an der Basis verschlossenen) Stacheln erheben. Die Form der Basischuppe ist deshalb schwer zu beurteilen und scheint mehr oder weniger rechteckig zu sein.
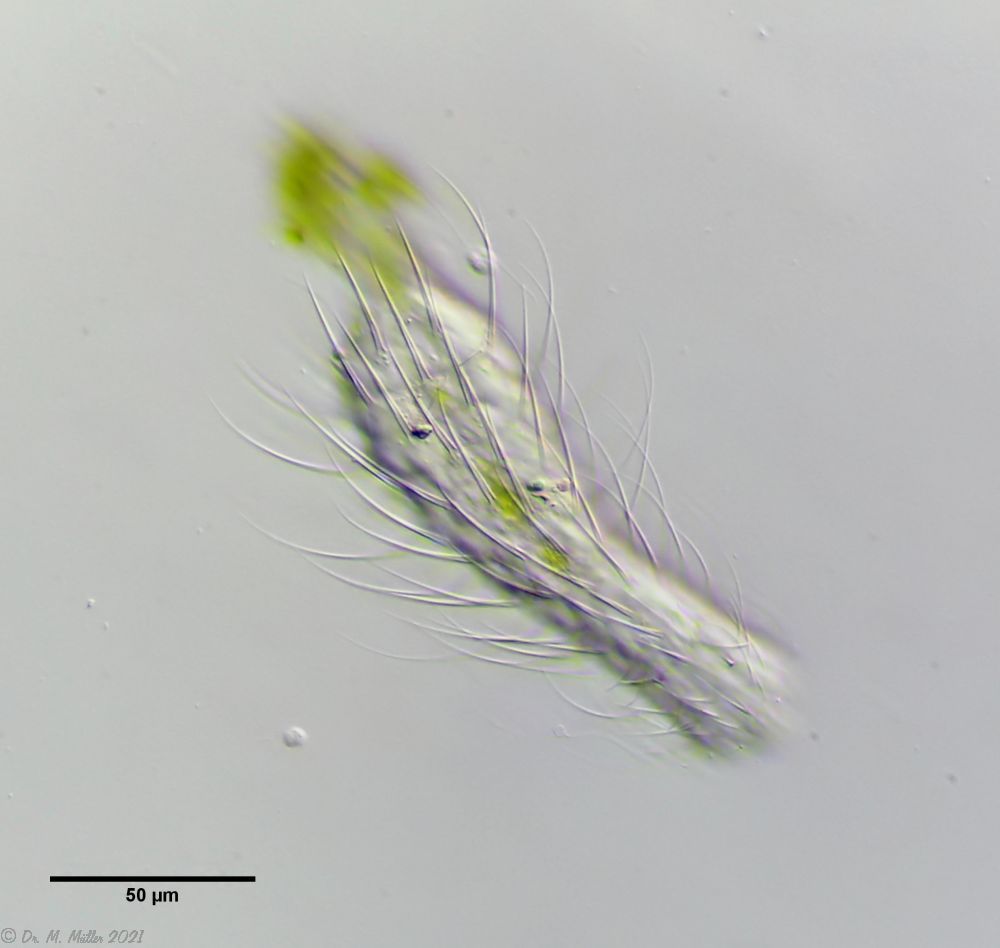
Bild 3: Ch. arquatus; dorsal
Die langen und dichten Stacheln verhindern ein gutes Portrait der Rückenseite, da hierfür die Tiefenschärfe nicht ausreicht (oder man das Tier unförmig platt drücken müsste). Dennoch erhält man einen ganz guten Eindruck davon, welche ambitionierte Beute dieses Tier für Räuber darstellen muss.
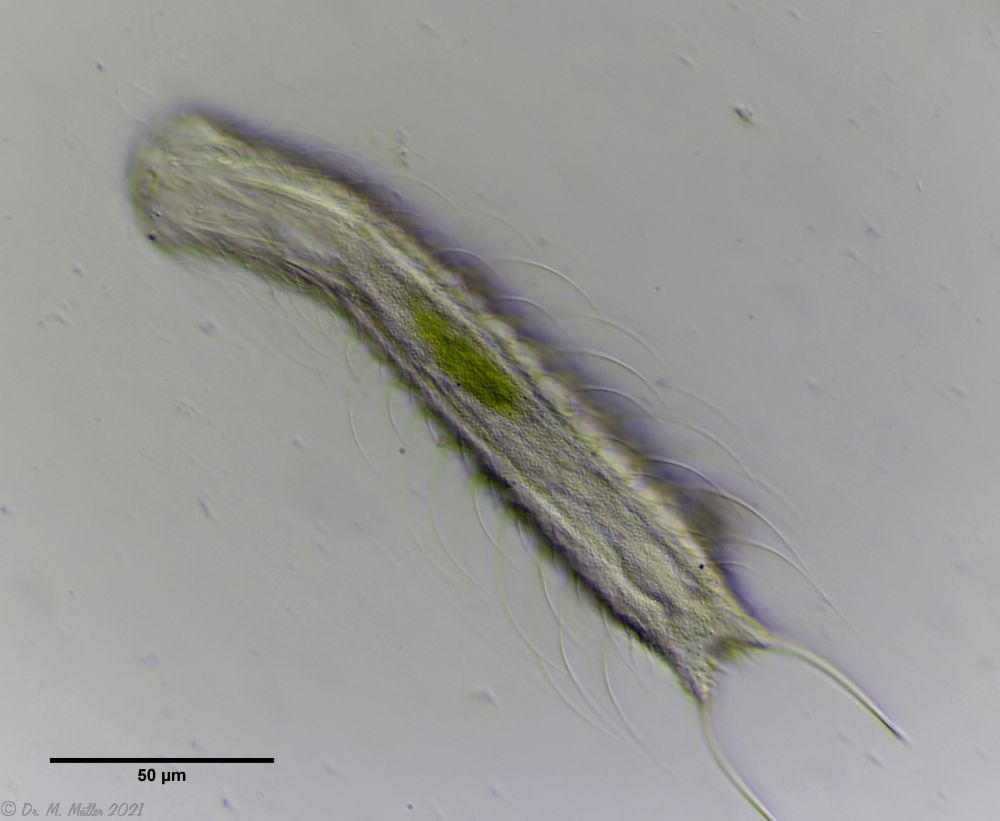
Bild 4: Ch. arquatus; ventral
Die Bauchseite aller von mir untersuchten, erwachsenen Exemplaren von Ch. arquatus ist durchgängig von einem Sekret unbekannter Funktion bedeckt, das besonders am Hinterende der Tiere auffällt. Dieses Sekret ist wasserlöslich, da es bei verstorbenen Tiere - wenn es nicht mehr nachgebildet wird - sich innerhalb weniger Stunden auflöst und die darunter verborgenen Schuppen freigibt. Bei Jungtieren ist diese Sekret noch nicht vorhanden. Hier kann die Schuppenform des ventralen Zwischenfeldes (des Feldes zwischen den beiden bauchseitigen Zilienbändern) beurteilt werden:
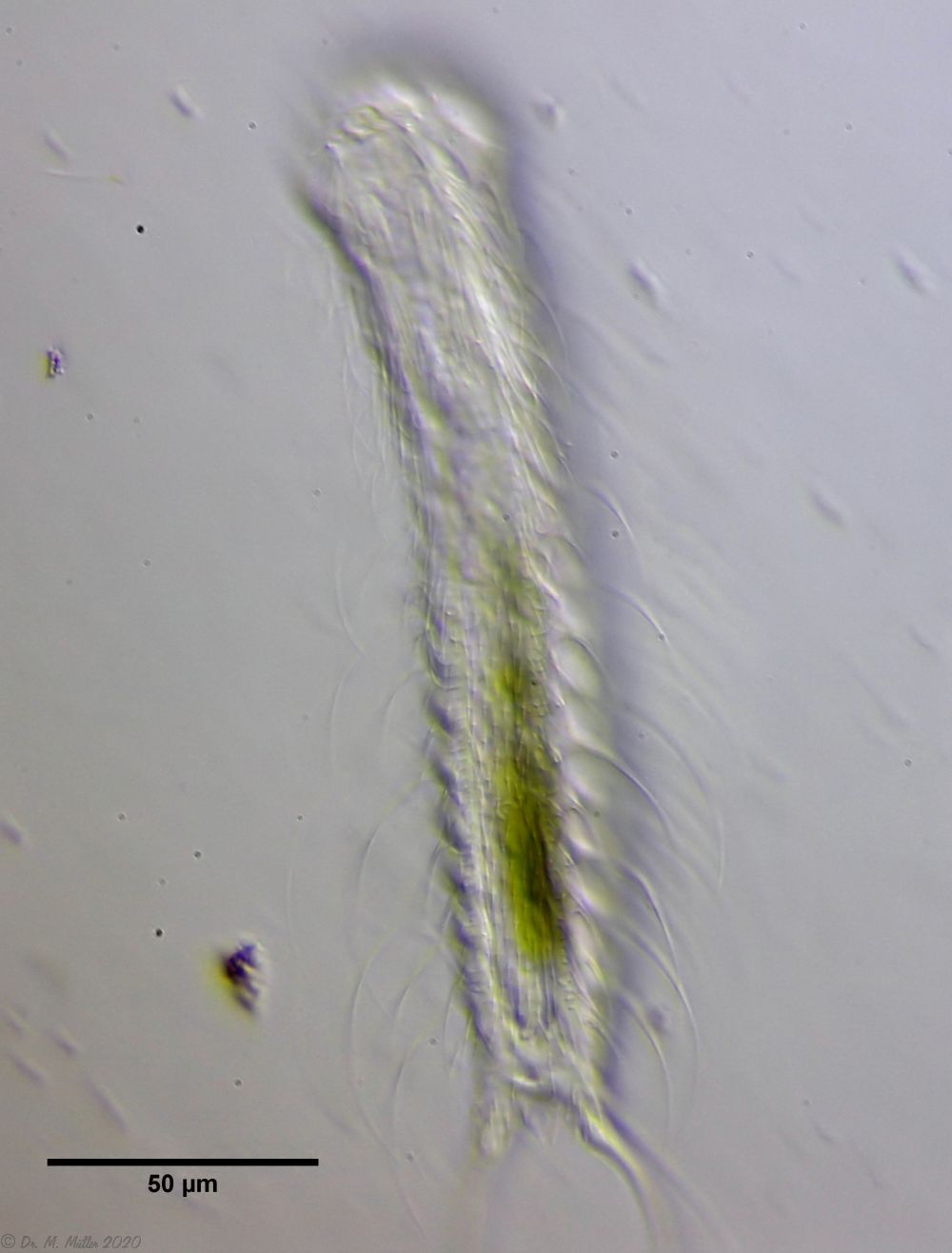
Bild 5: Ch. arquatus; Jungtier, Beschuppung des ventralen Zwischenfeldes
Hier findet man sieben Reihen sehr kleiner, länglicher Schuppen, die mit einem Mittelkiel und kleinen Endstacheln besetzt sind. Terminal erkennt man zwei etwas größere, ovale Kielschuppen.
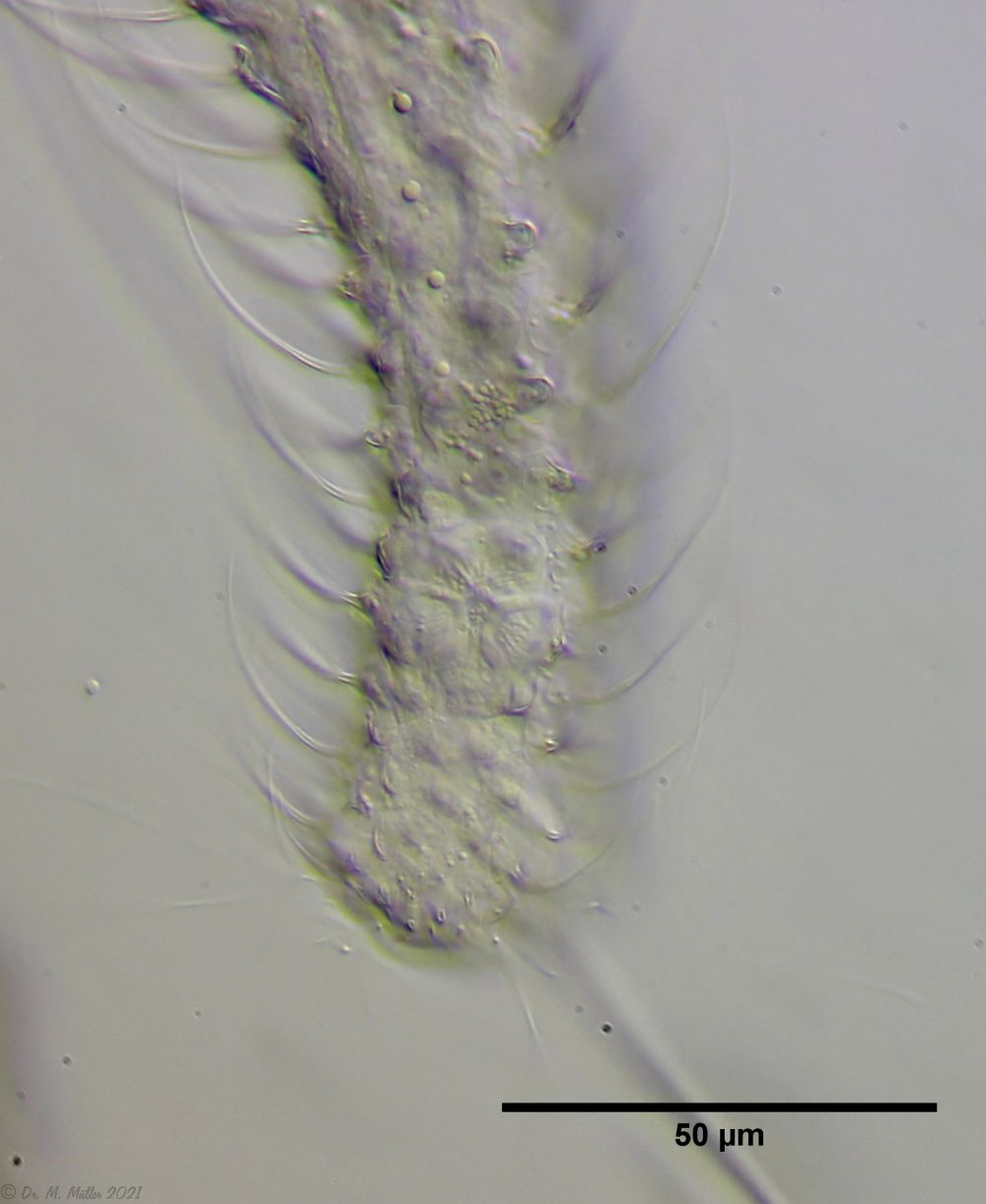
Bild 6: Ch. arquatus; Bulbus des Pharynx mit Gefäßen
Wie bei viele Gastrotrichen endet der Saugmagen (Pharynx) der Tiere in einen kugelförmigen Bulbus (Bild 1,2). Sehr deutlich erkennt man an der Oberfläche des Bulbus einige Gefäße, die auch bei anderen Gastrotrichen beschrieben sind, deren Funktion aber noch nicht geklärt ist.
Bis zu diesem Punkt entspricht die morphologische Beschreibung (abgesehen des Sekretfilmes auf der Bauchseite) von Ch. arquatus der eines zwar ungewöhnlichen Gastrotrichen, liegt aber durchaus innerhalb der normalen Variationsbreite für Gastrotrichen.
Vielleicht ist dem einen oder anderen Betrachter in den obigen Bilder ein eigenartiges Organ am Hinterende der Tiere aufgefallen:
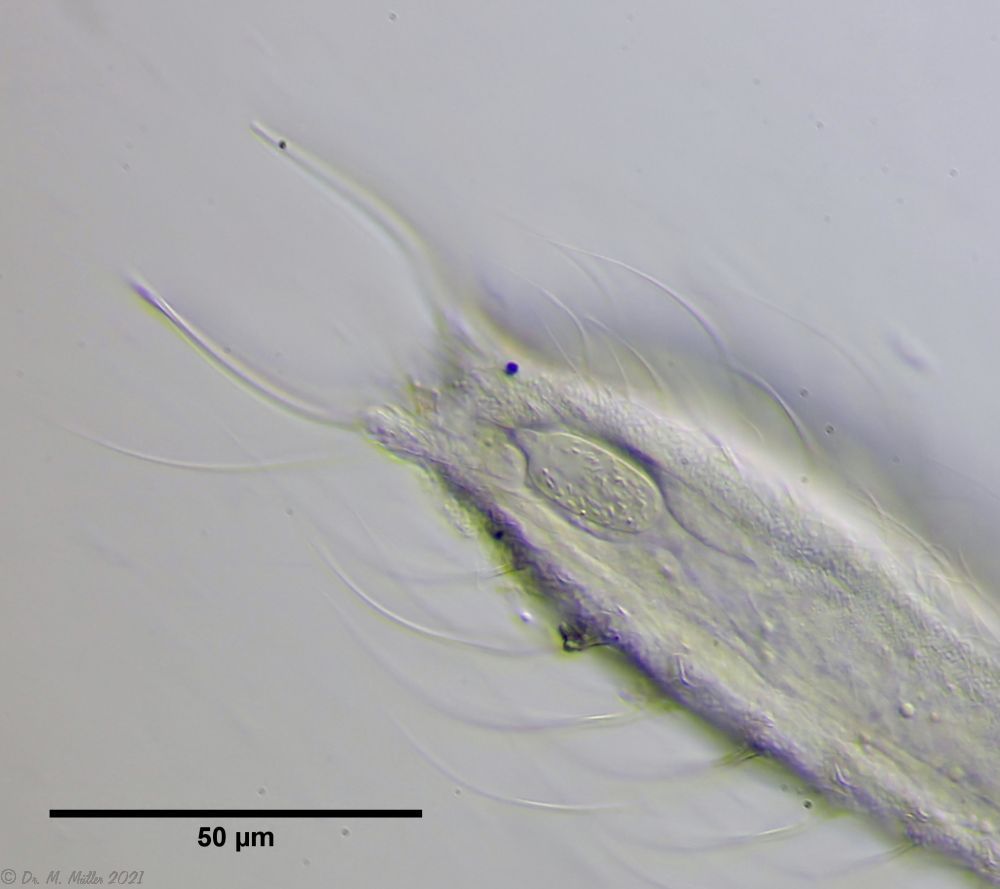
Bild 7: Ch. arquatus; caudales Organ
Bei allen untersuchten, erwachsenen Exemplaren fand sich unterhalb des Darms ein eigenartiges, ovoides Organ, das mit fädrigen, unbeweglichen Strukturen gefüllt ist und einen penisartigen Ausführungsgang auf die Ventralseite besitzt.
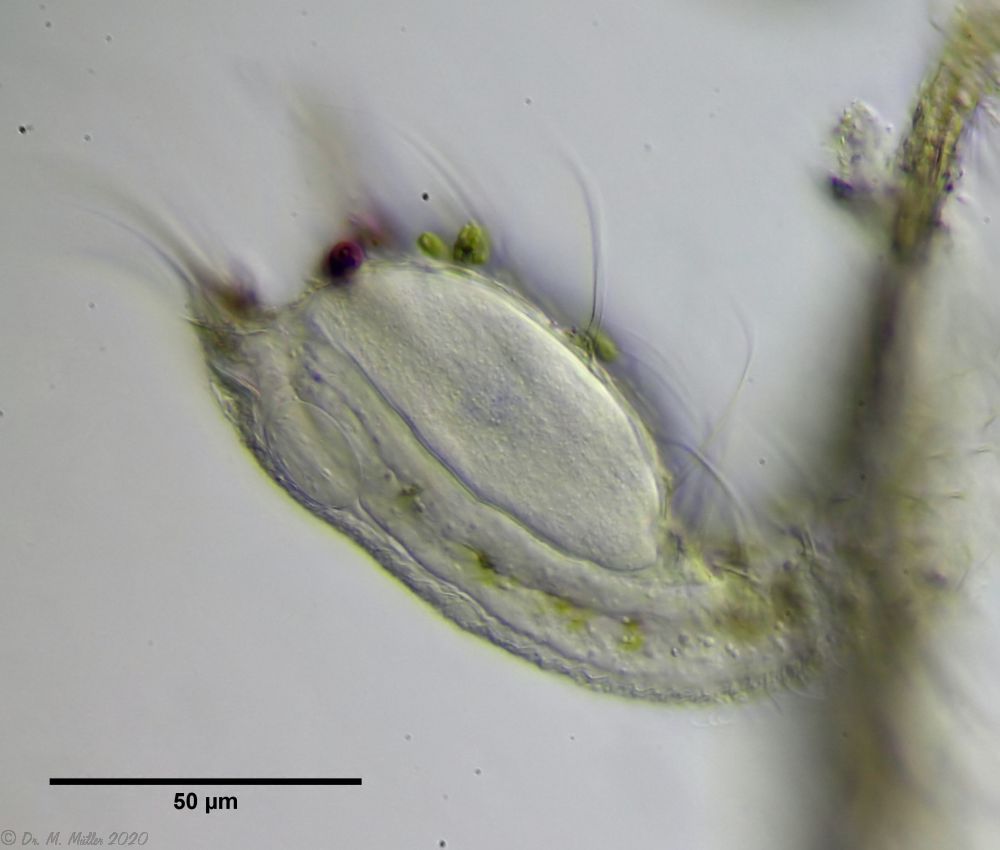
Bild 8: Ch. arquatus; caudales Organ, lateral
Dieses caudale Organ scheint von der Vordereite durch einen Zuführungsgang gespeist werden. An dem rückseitigem Ausführungsgang ist ein Ringmuskel zu erkennen, der diesen Gang nach Außen abschnürt:
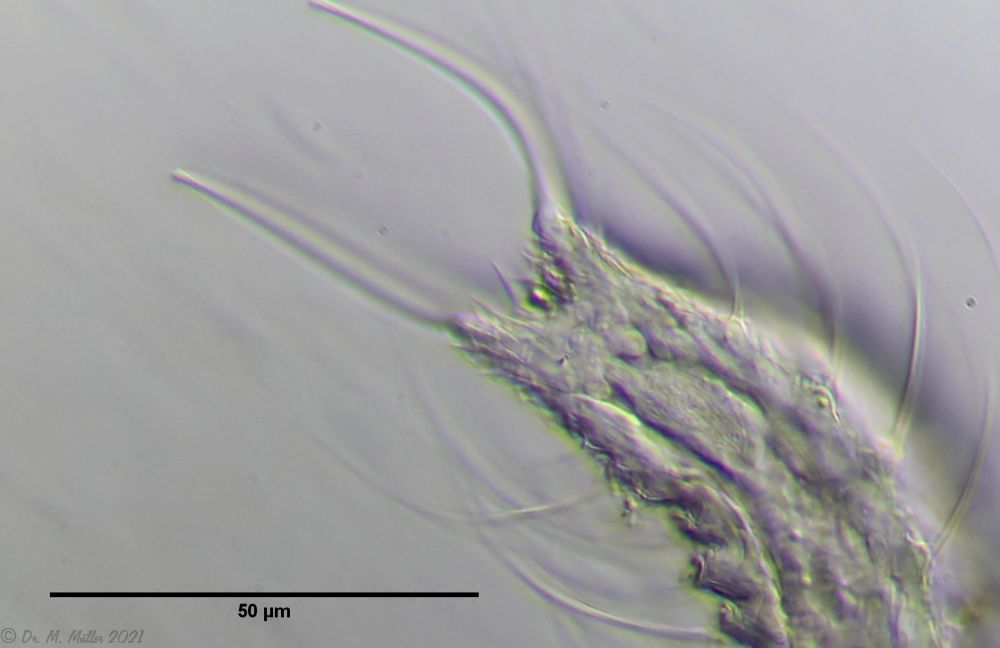
Bild 9: Ch. arquatus; caudales Organ mit Ringmuskel
Diese caudale Organ ist bei Jungtieren noch nicht (oder bestenfalls sehr undeutlich) zu sehen und bildet und füllt sich erst nach einigen Tagen:
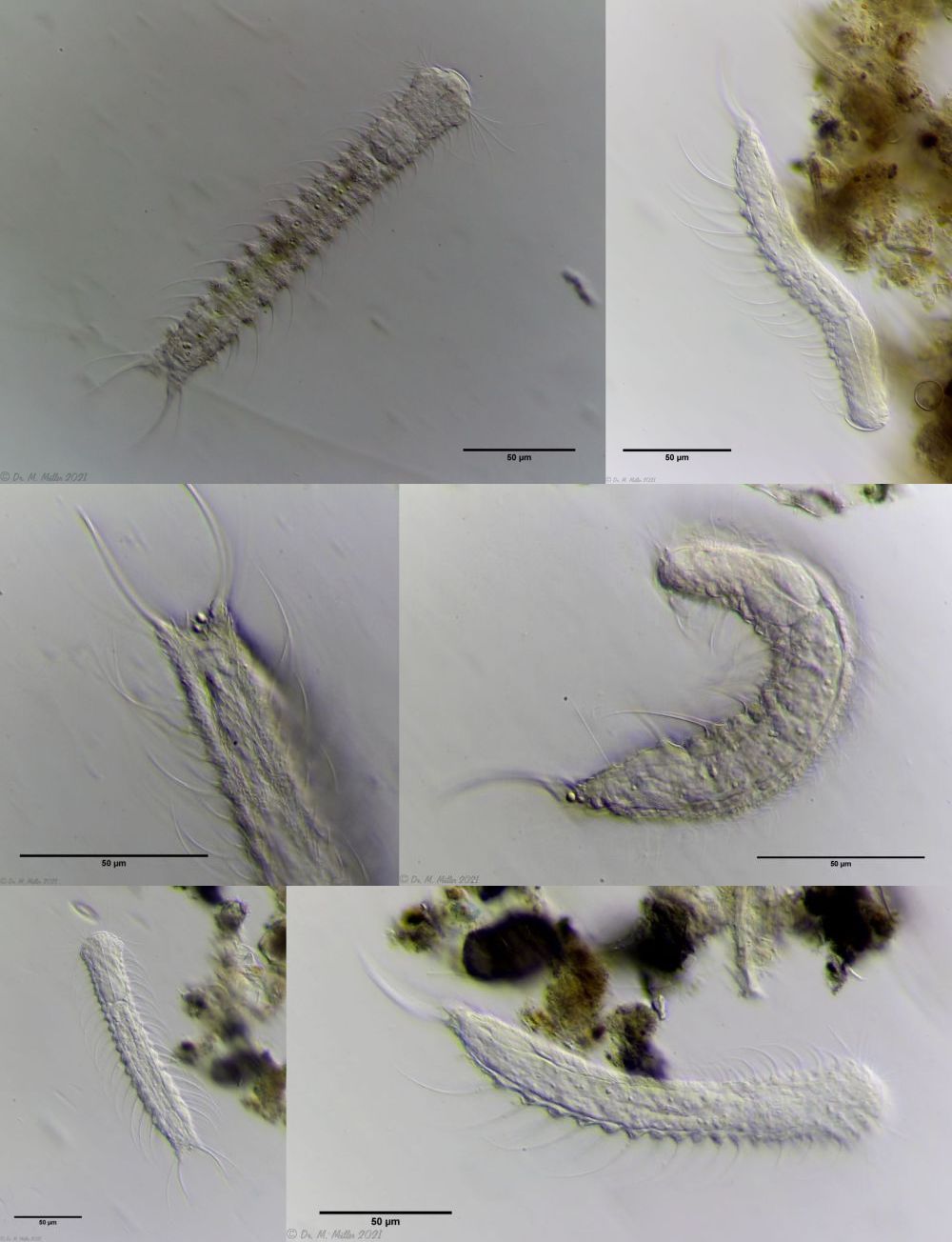
Bild 10: Ch. arquatus; Entwicklung des caudalen Organs im Abstand von jeweils 2 Tagen
Erst nach einigen Tagen ist die Entwicklung des caudalen Organs abgeschlossen:

Bild 11: Ch. arquatus; caudales Organ beim erwachsenen Tier
Ein ähnliches Organ habe ich bisher nur bei Ichthydium bifasciale gefunden ( https://www.mikroskopie-forum.de/index.php?topic=35723.0 ).
Bei einem Hohlorgan, dass mit fädrigen, unbeweglichen Strukturen gefüllt ist, liegt der Verdacht nahe, dass es sich um ein Speicherorgan für Sperma handelt. Ein solches Organ ist bei Süsswasser-Gastrotrichen nicht bekannt. Um diesen Verdacht zu überprüfen, habe ich versucht, den Organ-Inhalt mit Karmin-Essigsäure zu färben, die recht selektiv DNA färbt:
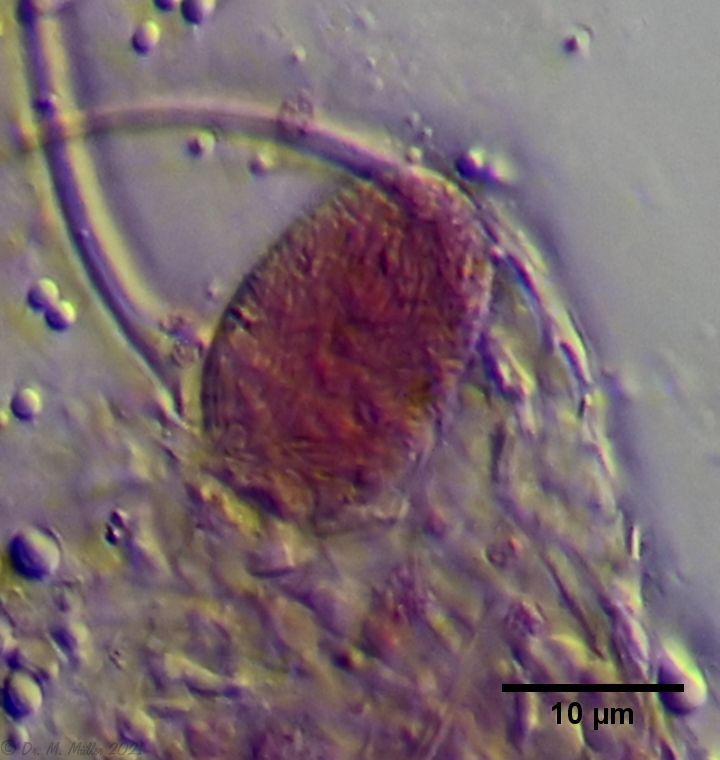
Bild 12: Ch. arquatus; caudales Organ mit Karmin-Essigsäure gefärbt
Auch wenn das Ergebnis der Färbung nicht 100% überzeugend ausfiel, wurde der Inhalt des caudalen Organs stark angefärbt. Das caudale Organ ist wohl mit DNA-haltigem, fädrigen Material angefüllt. Wenn das wirklich Sperma ist, wären die Tiere bereits als Jungtiere Zwitter. Dies unterscheidet sie von allen anderen bekannten Süsswasser-Gastrotrichen, die beim Schlupf rein parthenogenetische Weibchen sind und sich erst nach einigen Wochen langsam zu Zwittern entwickeln. Man geht davon aus, dass die parthenogenetische Fortpflanzung automiktisch, also durch diploide Eier erfolgt, die durch einfach mitotische Zellteilung entstanden sind. Die Nachkommen wären dann ein direkter Klon der Mutter. In der späteren Zwitterphase der Süsswasser-Gastrotrichen werden die Eier durch Meiose gebildet und sind haploid. Erst durch eine Verschmelzung mit einem anderen haloiden Gameten kommt es zur Bildung einer Zygote - der ersten somatischen Zelle des neuen Tieres.
Wenn Ch. arquatus bereits von Anfang an ein Zwitter ist, liegt es nahe, dass die reifen Eier durch Meiose gebildet wurden, so dass eine weitere Entwicklung nur nach der Verschmelzung mit einem zweiten haploiden Kern (z. B. durch eine Sperma-Zelle) erfolgen kann. Innerhalb meiner Mikroaquarien habe ich aber die Fortpflanzung isolierter Einzeltiere beobachtet. Diese Fortpflanzung erfolgte also entweder durch Selbstbefruchtung oder rein parthenogenetisch durch Automixis - der Verschmelzung zweier Zellen aus dem selben Meiose-Pfad. Es lag also nahe zu versuchen, den Beginn der Embryonalentwicklung nach der Eiablage genauer zu beobachten.
Ich verfolgte und filmte also ein Tier mit einem reifen Ei mehrere Stunden lang. Leider schaffte das Tier die Eibablage nicht und verstarb bei dem Versuch, das amitioniert große Ei abzulegen. Dies ist eine der häufigsten Todesursachen bei Gastrotrichen. In der Hoffnung, eine Wanderung von Spermatozoen zu beobachten, begann ich eine Zeitraffer-Aufnahme der toten Mutter.
Ich hatte Glück - das Ei begann seine Embryonalentwicklung trotz des Todes der Mutter - die Natur ist oft schrecklich effizient!
Im Folgenden möchte einige Standbilder des Films kommentieren. Dabei nehme ich an, dass das Ei durch eine Meiose entstanden ist und haploid ist (nur einen einfachen Chromosomensatz enthält). Die Kernkonfiguration vor der ersten Kernteilung und nach der DNA-Synthese ist also 1n2c (1n = einfacher Chromosomensatz; 2c = zwei Chromatiden, also gleiche Chromosomenäste, pro Chromosom).
Ausgangspunkt ist das einzellige Ei mit dem Zellkern, der einige auffällige Nucleoli enthällt.
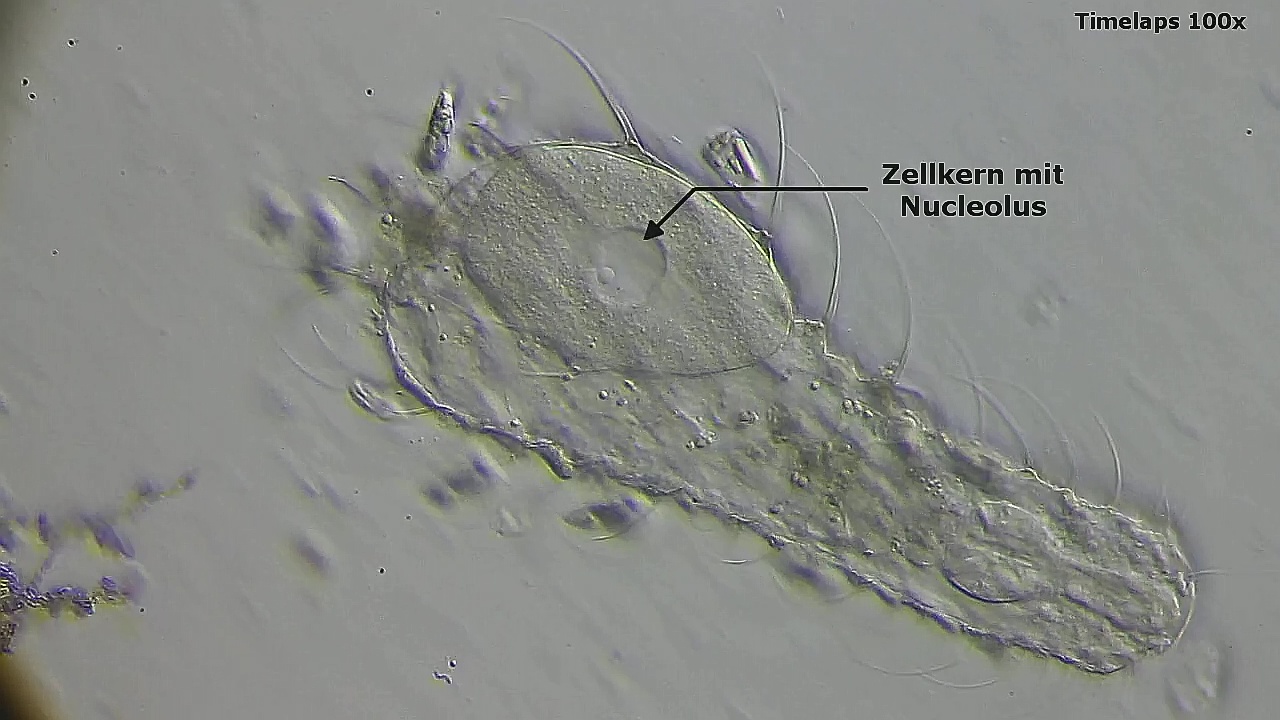
Bild 13: Ch. arquatus; Ei nach dem Tod der Mutter
Das Ei beginnt die weitere Entwicklung mit einer ersten Kernteilung:
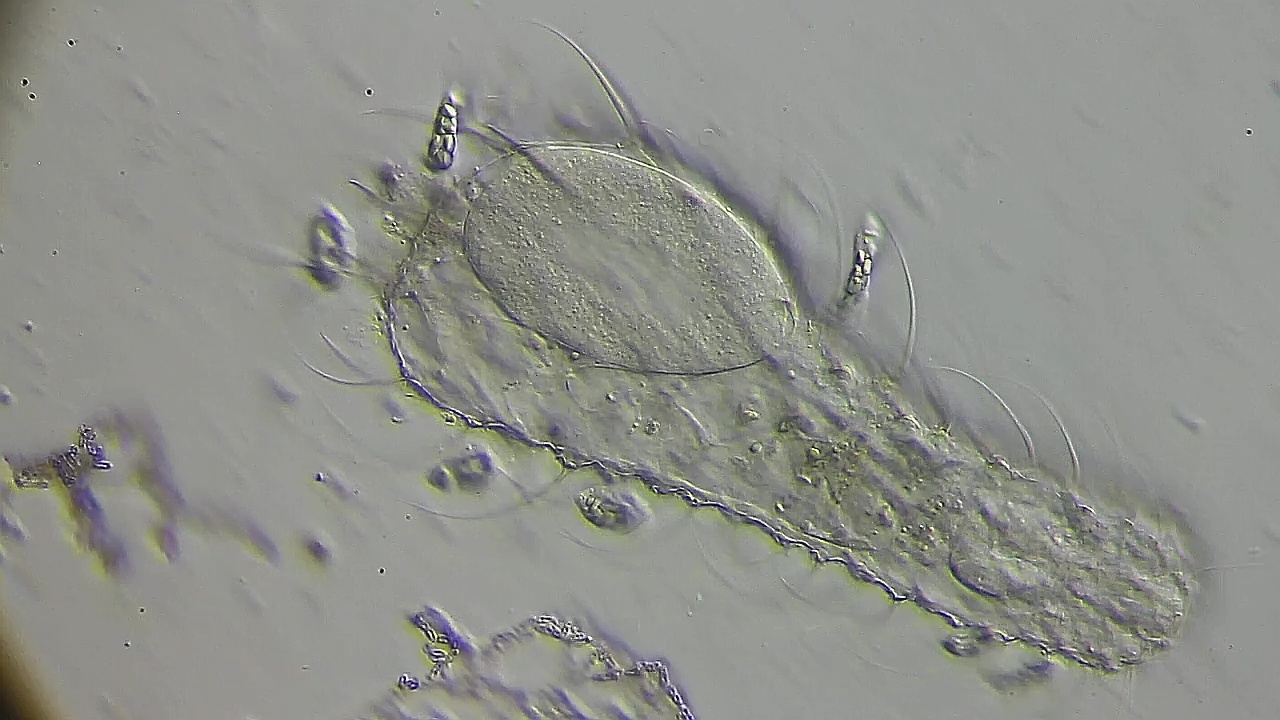
Bild 14: Ch. arquatus; Beginn der ersten Kernteilung: die Kernmembran löst sich auf
Es bildet sich eine Eizelle mit zwei Kernen; die Zelle teilt sich nicht. Die beiden 1n1C - Kerne beginnen in der Interphase mit der DNA-Synthese, die sie als 1n2c-Kerne abschließen:
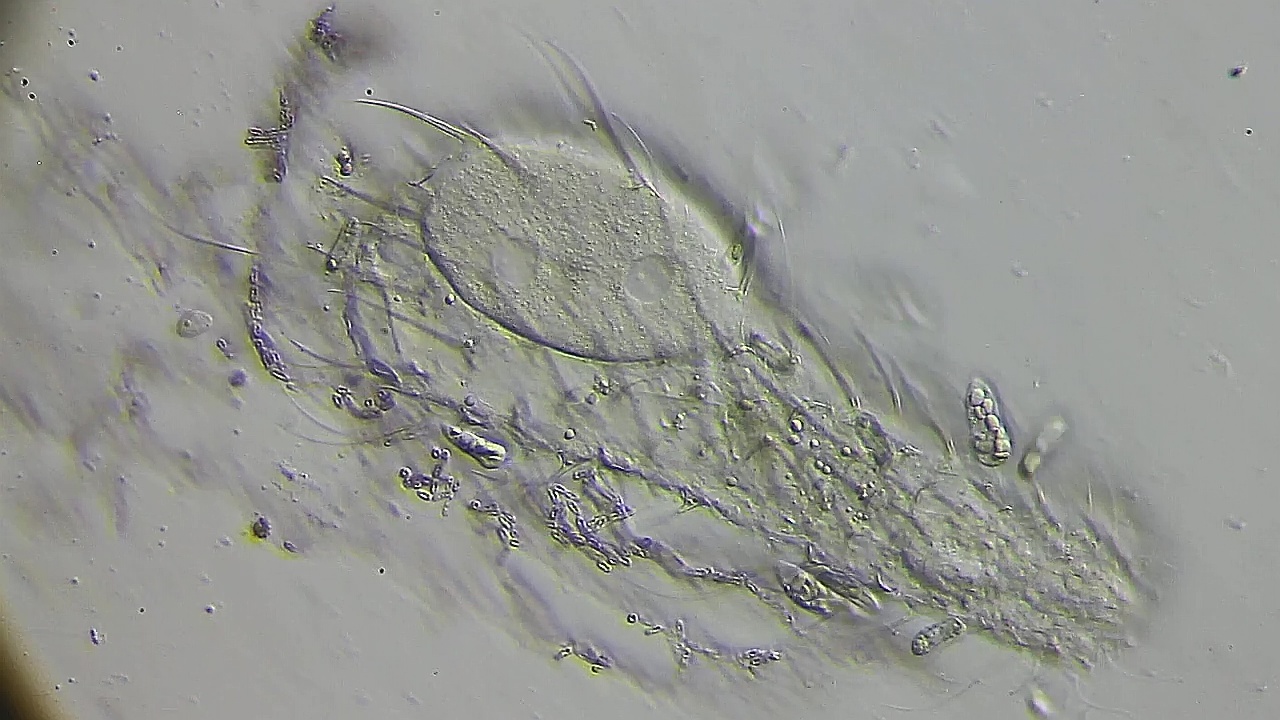
Bild 15: Ch. arquatus; eine Zelle mit zwei Interphasen-Kerne
Diese Kerne näher sich immer weiter an und verschmelzen zu einem 2n4c-Kern:
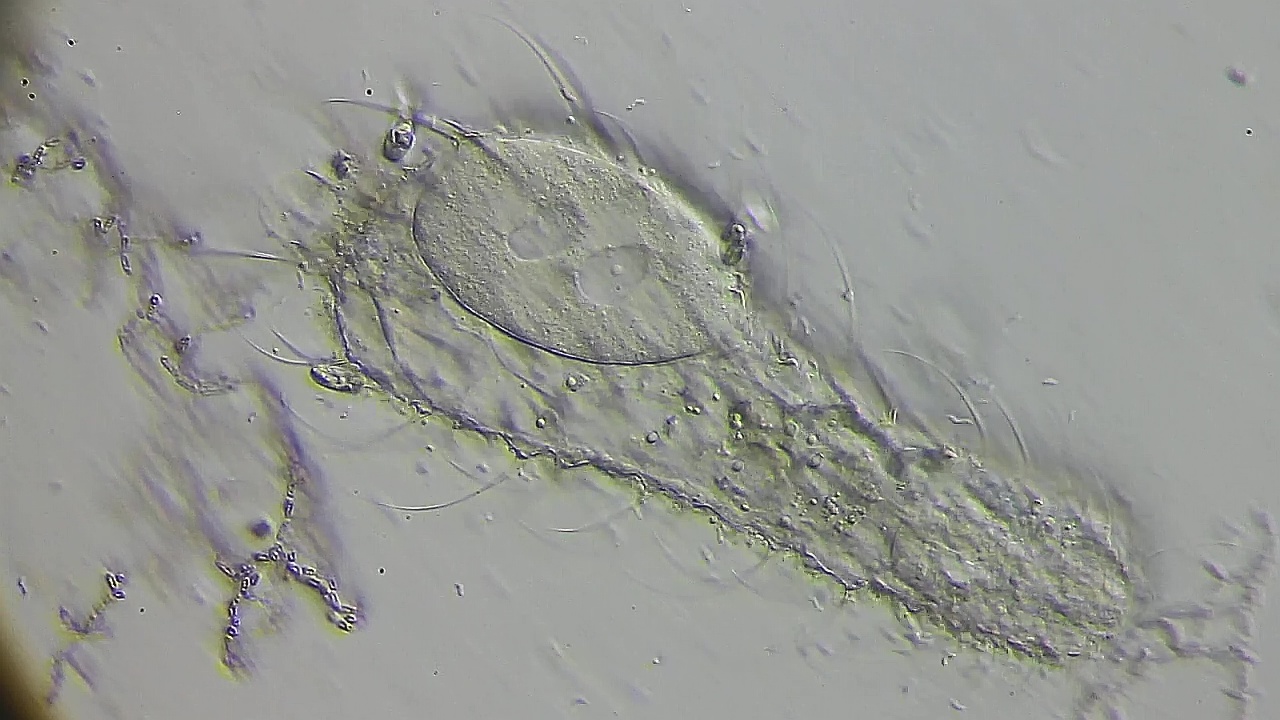
Bild 16: Ch. arquatus; kurz vor der Verschmelzung der haploiden Einzelkerne
Der neue Kern ist nun diploide und muss keine neue DNA vor der nächsten Teilung bilden. Deshalb tritt er nicht in eine Interphase ein und teilt sich sofort in die beiden ersten somatischen 2n2c-Kerne der neuen Tieres. Bei dieser Teilung lag der Fokus zufällig gerade richtig, um erstmals bei Gastrotrichen die Chromosomen bei der Kernteilung zu verfolgen.
Die Chromosomen ordnen sich während der Methaphase in der Äquatorialebene an:
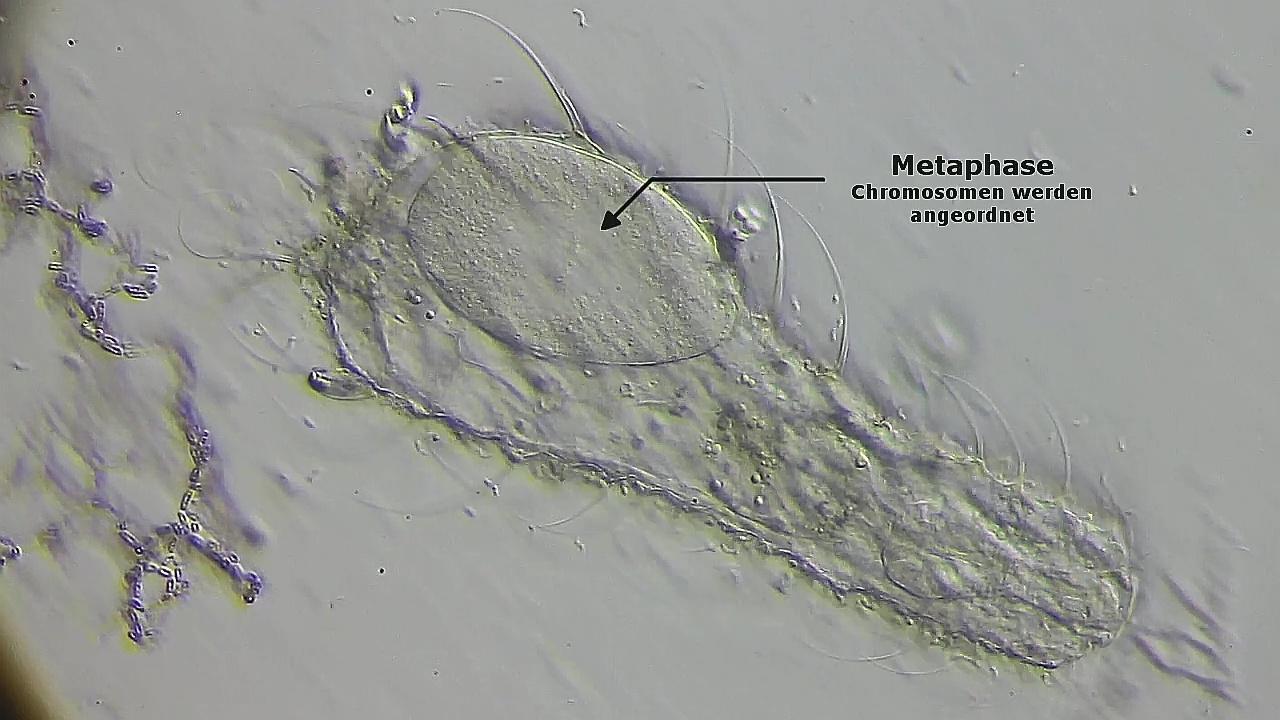
Bild 17: Ch. arquatus; Methaphase der Zellteilung; viele kleine Chromosomen sind direkt zu verfolgen
Nach einer leichten Drehung des Spindelkörpers werden sie Chromosomen während der Anaphase auseinander gezogen:
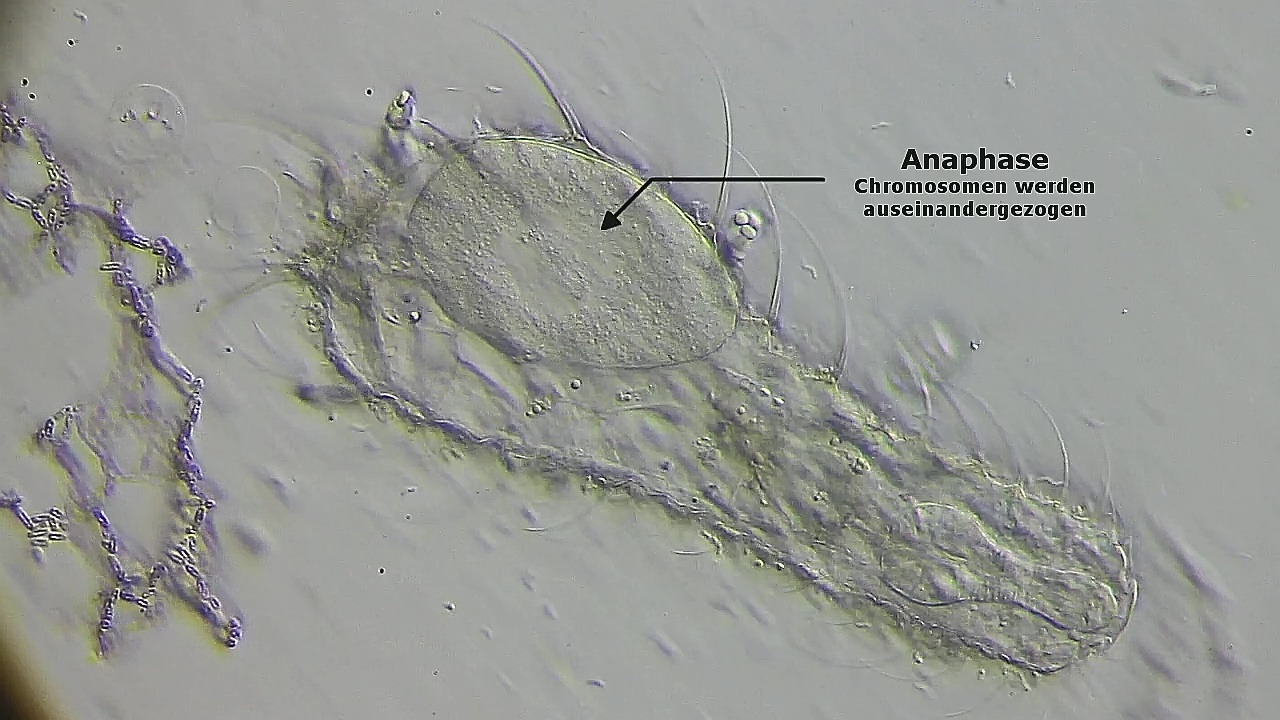
Bild 18: Ch. arquatus; Anaphase - die Chromosomen werden auseinander gezogen
In der Telophase dekondensieren die Chromosomen wieder und werden dadurch unsichtbar:
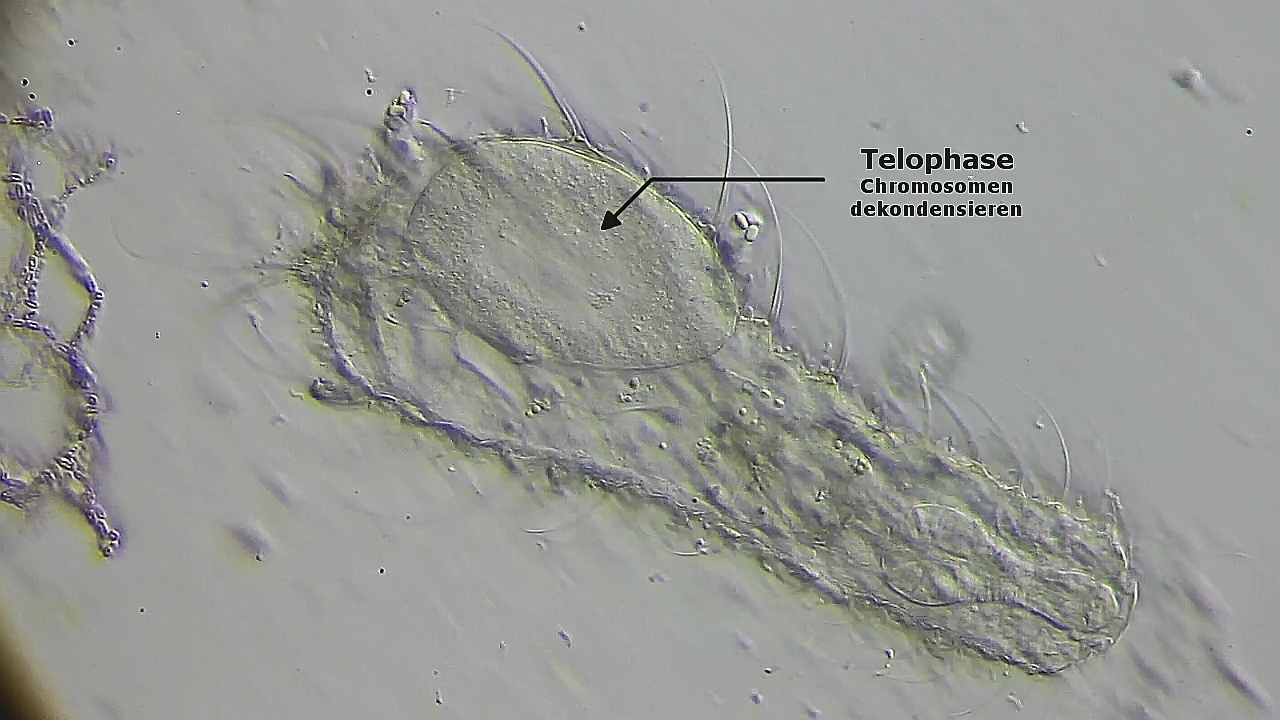
Bild 19: Ch. arquatus; Telophase; die Chromosomen dekondensieren
Die beiden Kerne beginnen ihre Kernmembran neu zu bilden. Leider verstarb das Ei ebenfalls zu diesem Zeitpunkt.
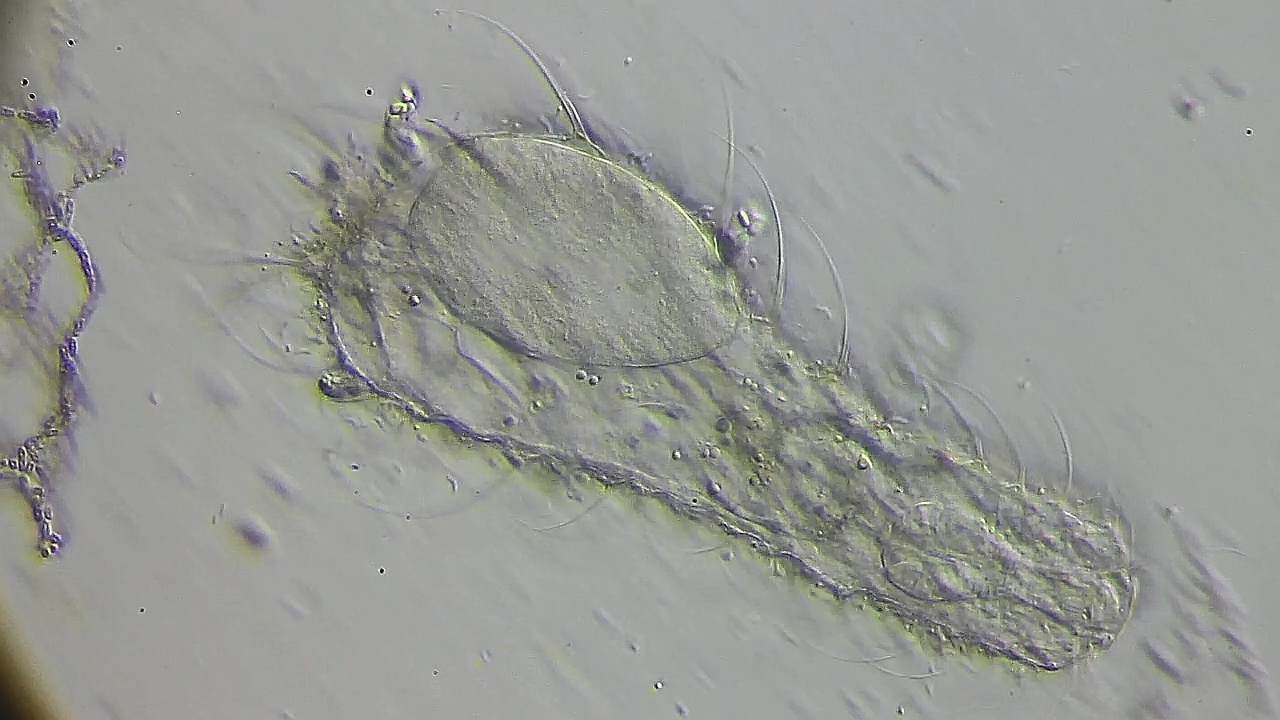
Bild 20: Ch. arquatus; Bildung der neuen Kernmembranen - Tod des Eies
Der gesamte Zeitraffer-Film kann hier angesehen werden:
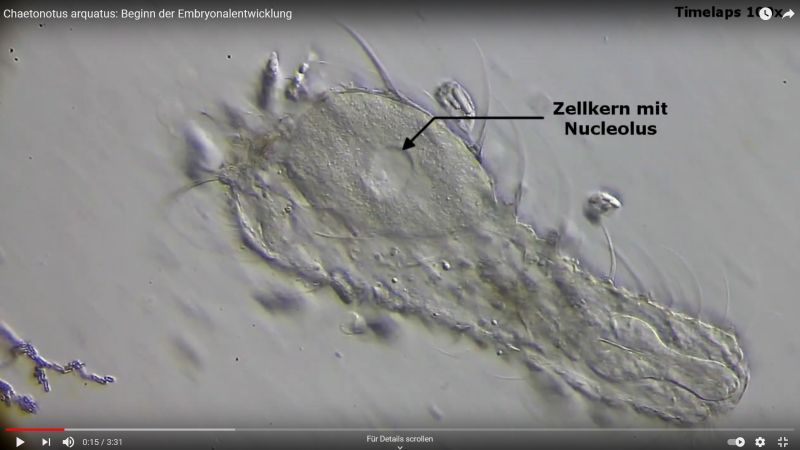
Bild 21: Ch. arquatus; Zum Starten bitte anklicken!
Der beobachtete Beginn der Embryonalentwicklung stützt also die Hypothese, dass Ch. arquatus haploide Eier bildet, die sich parthenogenetisch durch Automixis fortpflanzen. Dabei verschmelzen zwei haploide Kerne, die aus einer ersten Kernteilung des haploiden Eies hervorgehen. Eine Selbstbefruchtung konnte nicht beobachtet werden.
Der eigenartige Bauchhärling Chaetonotus arquatus scheint also nur als Zwitter zu existieren. Eine - wie bei den anderen Süsswassser-Gastrotrichen übliche - rein parthenogenetische Phase scheint nicht vor zukommen. Das stellt Ch. arquatus sehr isoliert innerhalb der Gastrotrichen, so dass eine genauere Abklärung der Gegebenheiten, als sie für mich als Amateur möglich ist, notwendig erscheint.
Bei meinen obigen Ausführungen handelt es sich sicherlich um Einzelbeobachtungen, die eine genauerer Überprüfung bedürfen. Dennoch kann ich mir die Beobachtungen nur durch die gegebenen Interpretationen erklären. Ich würde mich freuen, wenn Ihr andere Erklärungsversuche beisteuern könnt. Als fachfremder Amateur bin ich mir meiner Ausführungen oft nicht so sicher und für Korrekturen dankbar. Vielleicht kann das Schwarmwissen des Forums meine eigenartigen Beobachtungen besser einordnen.
Viele Grüße
Michael